

Antigenic conservation and immunogenicity of the HIV coreceptor binding site
- PMID: 15867093
- PMCID: PMC2213183
- DOI: 10.1084/jem.20042510
Antigenic conservation and immunogenicity of the HIV coreceptor binding site
Abstract
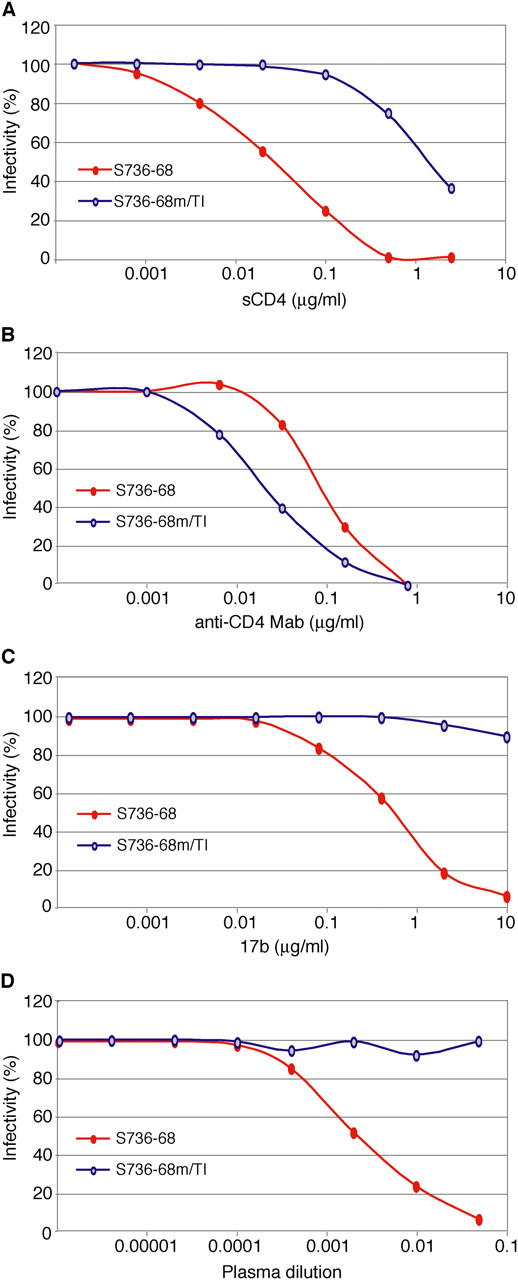
Immunogenic, broadly reactive epitopes of the HIV-1 envelope glycoprotein could serve as important targets of the adaptive humoral immune response in natural infection and, potentially, as components of an acquired immune deficiency syndrome vaccine. However, variability in exposed epitopes and a combination of highly effective envelope-cloaking strategies have made the identification of such epitopes problematic. Here, we show that the chemokine coreceptor binding site of HIV-1 from clade A, B, C, D, F, G, and H and circulating recombinant form (CRF)01, CRF02, and CRF11, elicits high titers of CD4-induced (CD4i) antibody during natural human infection and that these antibodies bind and neutralize viruses as divergent as HIV-2 in the presence of soluble CD4 (sCD4). 178 out of 189 (94%) HIV-1-infected patients had CD4i antibodies that neutralized sCD4-pretreated HIV-2 in titers (50% inhibitory concentration) as high as 1:143,000. CD4i monoclonal antibodies elicited by HIV-1 infection also neutralized HIV-2 pretreated with sCD4, and polyclonal antibodies from HIV-1-infected humans competed specifically with such monoclonal antibodies for binding. In vivo, variants of HIV-1 with spontaneously exposed coreceptor binding surfaces were detected in human plasma; these viruses were neutralized directly by CD4i antibodies. Despite remarkable evolutionary diversity among primate lentiviruses, functional constraints on receptor binding create opportunities for broad humoral immune recognition, which in turn serves to constrain the viral quasispecies.
Figures




References
-
- Albert, J., B. Abrahamsson, and K. Nagy. 1990. Rapid development of isolate-specific neutralizing antibodies after primary HIV-1 infection and consequent emergence of virus variants which resist neutralization by autologous sera. AIDS. 4:107–112. - PubMed
-
- Wei, X., J.M. Decker, S. Wang, H. Hui, J.C. Kappes, X. Wu, J.F. Salazar-Gonzalez, M.G. Salazar, J.M. Kilby, M.S. Saag, et al. 2003. Antibody neutralization and escape by HIV-1. Nature. 422:307–312. - PubMed
-
- Pilgrim, A.K., G. Pantaleo, O.J. Cohen, L.M. Fink, J.Y. Zhou, J.T. Zhou, D.P. Bolognesi, A.S. Fauci, and D.C. Montefiori. 1997. Neutralizing antibody responses to human immunodeficiency virus type 1 in primary infection and long-term-nonprogressive infection. J. Infect. Dis. 176:924–932. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
