

Review
doi: 10.1146/annurev.biophys.34.040204.144534.
Use of EPR power saturation to analyze the membrane-docking geometries of peripheral proteins: applications to C2 domains
Affiliations
- PMID: 15869384
- PMCID: PMC3637887
- DOI: 10.1146/annurev.biophys.34.040204.144534
Item in Clipboard
Review
Use of EPR power saturation to analyze the membrane-docking geometries of peripheral proteins: applications to C2 domains
Annu Rev Biophys Biomol Struct.
2005.
Abstract
Despite the central importance of peripheral membrane proteins to cellular signaling and metabolic pathways, the structures of protein-membrane interfaces remain largely inaccessible to high-resolution structural methods. In recent years a number of laboratories have contributed to the development of an electron paramagnetic resonance (EPR) power saturation approach that utilizes site-directed spin labeling to determine the key geometric parameters of membrane-docked proteins, including their penetration depths and angular orientations relative to the membrane surface. Representative applications to Ca(2+)-activated, membrane-docking C2 domains are described.
Figures

Use of site-directed spin labeling and power saturation to measure depth parameters (, –15, 19, 23, 24). (a) Structure of the MTSSL nitroxide probe used for site-directed spin labeling, illustrating its coupling to a cysteine via a disulfide linkage. (b) The two gradients of paramagnetic probes used to define the depth parameter. Molecular oxygen is found at highest concentration in the bilayer hydrocarbon core because of its a polarity: Its concentration gradient points toward the membrane center. The zwitterionic nickel(II) ethylenediaminediacetic acid (NiEDDA) complex is found at highest concentration in the aqueous phase because of its high polarity: Its concentration gradient points away from the membrane. (c) Continuous-wave power saturation curves for a membrane-docked cysteine variant of cPLA2 C2 domain (Cys38-MTSSL) (24). The EPR signal amplitude is plotted as a function of the square root of microwave power. Saturation curves are shown for measurements of samples purged with nitrogen gas (closed circles), samples equilibrated with atmospheric oxygen (open circles), and samples containing 10 mM NiEDDA and purged with nitrogen gas (closed squares).

Plot of measured depth parameters versus modeled membrane depth for a membrane-docking C2 domain (24). The depth parameters of spin labels incorporated into the cPLA2 C2 domain are plotted as a function of the modeled distance from the headgroup phosphate plane. Filled circles indicate spin label positions in the membrane interior where the depth parameter varies linearly: These depth parameters were used in the linear fitting procedure that generated the membrane-docking model. Open circles indicate spin label positions outside the linear region that were excluded from the fitting procedure. The side chain conformations of six spin labels were adjusted to optimize the linear fit. Open squares indicate the measured depth parameters for phosphatidylcholine lipids possessing spin labels at different carbons in the A2 fatty acid, plotted as a function of the known distance of the fatty acid carbon from the plane of membrane phosphates (squares). The solid curve represents the best-fit hyperbolic function (Equation 4), illustrating the ability of this function to accurately reproduce the distance-dependence of the depth parameter.

Orientation and depth of the C2 domain of cPLA2 with respect to a membrane surface (24). The crystal structure of the C2 domain of cPLA2 (31) is represented as the cyan ribbon, with two Ca2+ ions shown as yellow spheres. The horizontal lines represent the planar boundaries of the headgroup and hydrocarbon regions of the bilayer. Protein spin labels oriented in their final optimized conformations are colored according to their measured depth parameters, with positive depth parameters indicated by increasing red, and negative depth parameters indicated by increasing blue. Figure generated by Insight (Accelrys).

Comparison of the membrane-docking geometries of the cPLA2, PKCα, and SytIA C2 domains (14, 15, 19, 23, 24). The cPLA2 C2 domain penetrates significantly farther into the hydrocarbon core, explaining its hydrophobic docking mechanism, and the PKCα and SytIA C2 domains interact primarily with the charged headgroup region, explaining their electrostatic docking mechanism. The cPLA2 C2 domain is oriented with its β-strands closer to the membrane normal than the other two domains. For all three domains the Ca2+ ions lie near the headgroup phosphate plane. Figure was generated by MacPymol (DeLano Scientific).
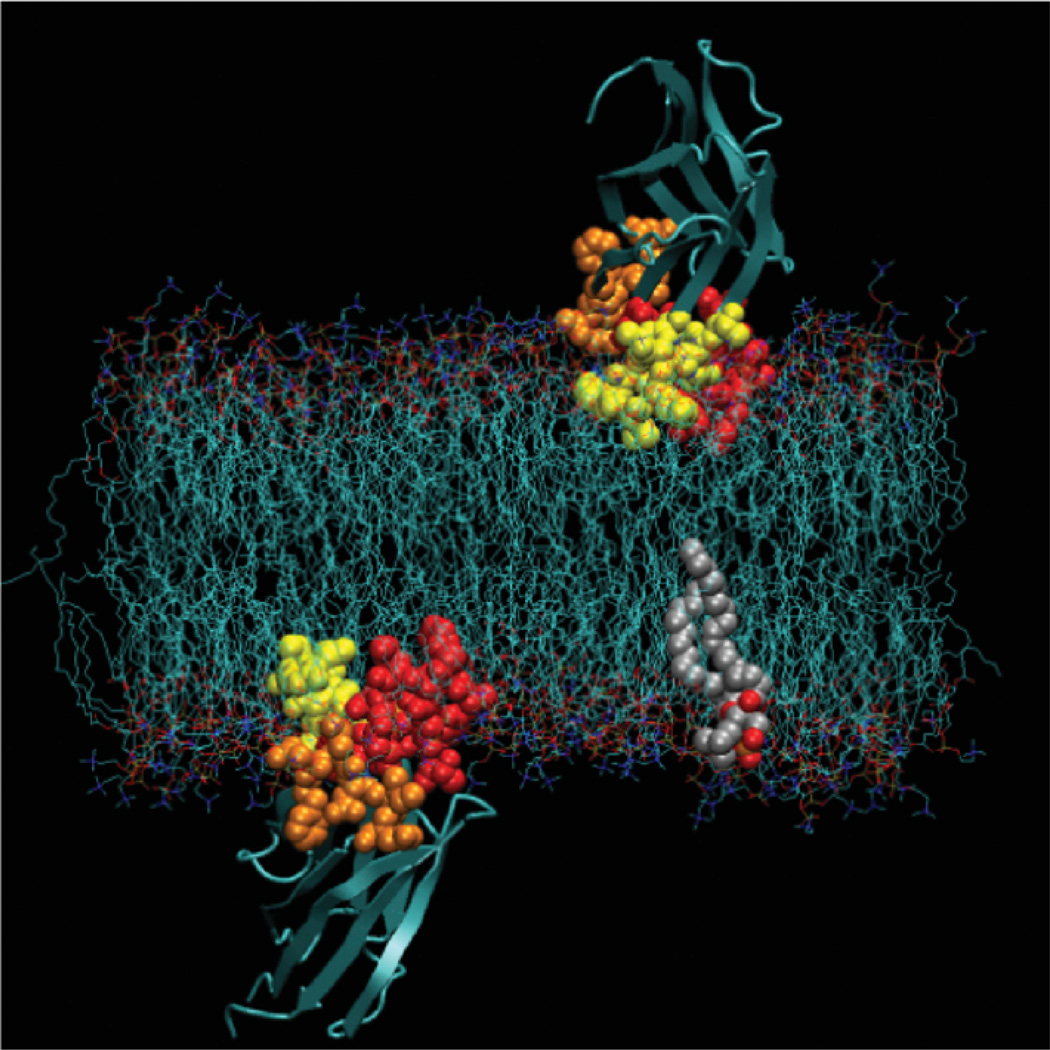
Molecular model of cPLA2 C2 domain docked to a lipid bilayer showing two views of the domain in the preferred docking geometry. The three Ca2+-binding loops are shown in CPK format with loop 1 in red, loop 2 in orange, and loop 3 in yellow. Also highlighted in CPK is a representative phospholipid (S. Jaud, S. White, J. Falke & D. Tobias, unpublished data). The model was generated by equilibrating the C2 domain in the membrane at the depth and angular orientation specified by the EPR membrane depth analysis (24), followed by unconstrained molecular dynamics that revealed that the system was stable for the full length of the molecular dynamics simulation (10 ns; see text).
Similar articles
-
Effect of PIP2 binding on the membrane docking geometry of PKC alpha C2 domain: an EPR site-directed spin-labeling and relaxation study.Biochemistry. 2008 Aug 12;47(32):8301-16. doi: 10.1021/bi800711t. Epub 2008 Jul 9. Biochemistry. 2008. PMID: 18610985 Free PMC article.
-
C2 domain of protein kinase C alpha: elucidation of the membrane docking surface by site-directed fluorescence and spin labeling.Biochemistry. 2003 Feb 11;42(5):1254-65. doi: 10.1021/bi026596f. Biochemistry. 2003. PMID: 12564928 Free PMC article.
-
Membrane-docking loops of the cPLA2 C2 domain: detailed structural analysis of the protein-membrane interface via site-directed spin-labeling.Biochemistry. 2003 Nov 18;42(45):13227-40. doi: 10.1021/bi035119+. Biochemistry. 2003. PMID: 14609334 Free PMC article.
-
High-field EPR on membrane proteins - crossing the gap to NMR.Prog Nucl Magn Reson Spectrosc. 2013 Nov;75:1-49. doi: 10.1016/j.pnmrs.2013.07.002. Epub 2013 Jul 29. Prog Nucl Magn Reson Spectrosc. 2013. PMID: 24160760 Review.
-
Attaching a spin to a protein -- site-directed spin labeling in structural biology.Acta Biochim Pol. 2007;54(2):235-44. Epub 2007 Jun 14. Acta Biochim Pol. 2007. PMID: 17565387 Review.
Cited by
-
The cellular membrane as a mediator for small molecule interaction with membrane proteins.Biochim Biophys Acta. 2016 Oct;1858(10):2290-2304. doi: 10.1016/j.bbamem.2016.04.016. Epub 2016 May 6. Biochim Biophys Acta. 2016. PMID: 27163493 Free PMC article.
-
Mechanism of specific membrane targeting by C2 domains: localized pools of target lipids enhance Ca2+ affinity.Biochemistry. 2007 Apr 10;46(14):4322-36. doi: 10.1021/bi062140c. Epub 2007 Mar 17. Biochemistry. 2007. PMID: 17367165 Free PMC article.
-
Effect of PIP2 binding on the membrane docking geometry of PKC alpha C2 domain: an EPR site-directed spin-labeling and relaxation study.Biochemistry. 2008 Aug 12;47(32):8301-16. doi: 10.1021/bi800711t. Epub 2008 Jul 9. Biochemistry. 2008. PMID: 18610985 Free PMC article.
-
Differential Membrane Binding Mechanics of Synaptotagmin Isoforms Observed in Atomic Detail.Biochemistry. 2017 Jan 10;56(1):281-293. doi: 10.1021/acs.biochem.6b00468. Epub 2016 Dec 20. Biochemistry. 2017. PMID: 27997124 Free PMC article.
-
Electrostatic interactions and binding orientation of HIV-1 matrix studied by neutron reflectivity.Biophys J. 2010 Oct 20;99(8):2516-24. doi: 10.1016/j.bpj.2010.07.062. Biophys J. 2010. PMID: 20959092 Free PMC article.
References
-
- Altenbach C, Flitsch SL, Khorana HG, Hubbell WL. Structural studies on transmembrane proteins. 2. Spin labeling of bacteriorhodopsin mutants at unique cysteines. Biochemistry. 1989;28:7806–7812. - PubMed
-
- Altenbach C, Oh KJ, Trabanino RJ, Hideg K, Hubbell WL. Estimation of interresidue distances in spin labeled proteins at physiological temperatures: experimental strategies and practical limitations. Biochemistry. 2001;40:15471–15482. - PubMed
-
- Bai J, Tucker WC, Chapman ER. PIP2 increases the speed of response of synaptotagmin and steers its membrane-penetration activity toward the plasma membrane. Nat. Struct. Mol. Biol. 2004;11:36–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous