

Role of epidermal growth factor receptor signaling in RAS-driven melanoma
- PMID: 15870287
- PMCID: PMC1087708
- DOI: 10.1128/MCB.25.10.4176-4188.2005
Role of epidermal growth factor receptor signaling in RAS-driven melanoma
Abstract
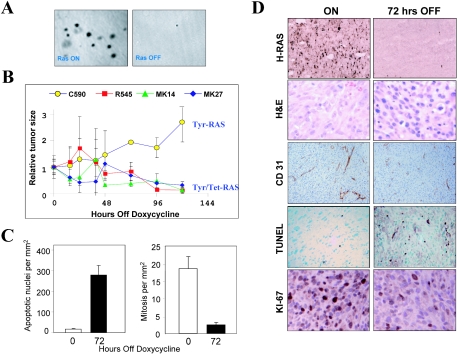
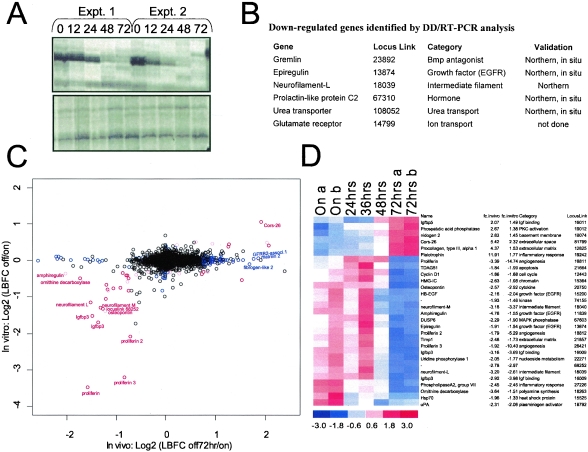
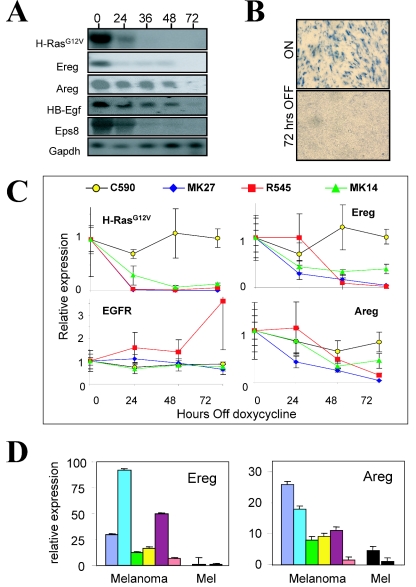
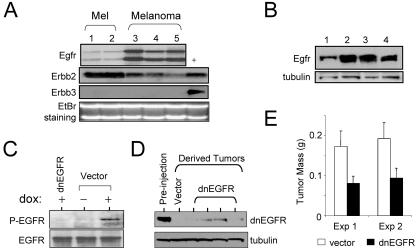
The identification of essential genetic elements in pathways governing the maintenance of fully established tumors is critical to the development of effective antioncologic agents. Previous studies revealed an essential role for H-RAS(V12G) in melanoma maintenance in an inducible transgenic model. Here, we sought to define the molecular basis for RAS-dependent tumor maintenance through determination of the H-RAS(V12G)-directed transcriptional program and subsequent functional validation of potential signaling surrogates. The extinction of H-RAS(V12G) expression in established tumors was associated with alterations in the expression of proliferative, antiapoptotic, and angiogenic genes, a profile consistent with the observed phenotype of tumor cell proliferative arrest and death and endothelial cell apoptosis during tumor regression. In particular, these melanomas displayed a prominent RAS-dependent regulation of the epidermal growth factor (EGF) family, leading to establishment of an EGF receptor signaling loop. Genetic complementation and interference studies demonstrated that this signaling loop is essential to H-RAS(V12G)-directed tumorigenesis. Thus, this inducible tumor model system permits the identification and validation of alternative points of therapeutic intervention without neutralization of the primary genetic lesion.
Figures
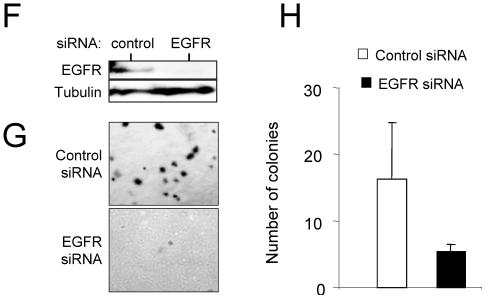
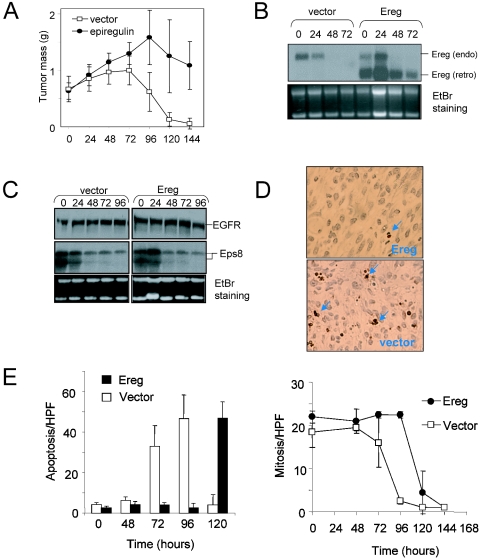
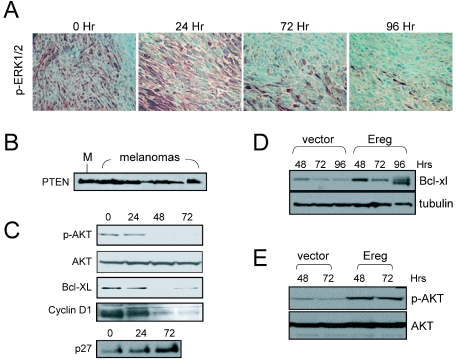
Similar articles
-
Ras-induced melanoma transformation is associated with the proteasomal degradation of the transcriptional repressor ICER.Mol Carcinog. 2013 Sep;52(9):692-704. doi: 10.1002/mc.21908. Epub 2012 Apr 4. Mol Carcinog. 2013. PMID: 22488648
-
Opposing roles of the extracellular signal-regulated kinase and p38 mitogen-activated protein kinase cascades in Ras-mediated downregulation of tropomyosin.Mol Cell Biol. 2002 Apr;22(7):2304-17. doi: 10.1128/MCB.22.7.2304-2317.2002. Mol Cell Biol. 2002. PMID: 11884615 Free PMC article.
-
TDAG51 is an ERK signaling target that opposes ERK-mediated HME16C mammary epithelial cell transformation.BMC Cancer. 2008 Jul 2;8:189. doi: 10.1186/1471-2407-8-189. BMC Cancer. 2008. PMID: 18597688 Free PMC article.
-
Molecular pathogenesis of cutaneous melanocytic neoplasms.Annu Rev Pathol. 2009;4:551-79. doi: 10.1146/annurev.pathol.3.121806.151541. Annu Rev Pathol. 2009. PMID: 19400696 Review.
-
Oncogenic ras-induced expression of cytokines: a new target of anti-cancer therapeutics.Mol Interv. 2008 Feb;8(1):22-7. doi: 10.1124/mi.8.1.6. Mol Interv. 2008. PMID: 18332481 Free PMC article. Review.
Cited by
-
WIPI1 promotes fission of endosomal transport carriers and formation of autophagosomes through distinct mechanisms.Autophagy. 2021 Nov;17(11):3644-3670. doi: 10.1080/15548627.2021.1886830. Epub 2021 Mar 8. Autophagy. 2021. PMID: 33685363 Free PMC article.
-
Guidelines for biomarker testing in metastatic melanoma: a National Consensus of the Spanish Society of Pathology and the Spanish Society of Medical Oncology.Clin Transl Oncol. 2014 Apr;16(4):362-73. doi: 10.1007/s12094-013-1090-5. Epub 2013 Oct 16. Clin Transl Oncol. 2014. PMID: 24129426
-
Effects of somatic alterations at pathway level are more mechanism-explanatory and clinically applicable to quantity of liver metastases of colorectal cancer.Cancer Med. 2019 Aug;8(10):4732-4742. doi: 10.1002/cam4.2368. Epub 2019 Jun 20. Cancer Med. 2019. PMID: 31219228 Free PMC article.
-
Drug targets and predictive biomarkers in the management of metastatic melanoma.Pharmgenomics Pers Med. 2012;5:139-48. doi: 10.2147/PGPM.S25100. Epub 2012 Sep 28. Pharmgenomics Pers Med. 2012. PMID: 23226069 Free PMC article.
-
EGFRAP encodes a new negative regulator of the EGFR acting in both normal and oncogenic EGFR/Ras-driven tissue morphogenesis.PLoS Genet. 2021 Aug 19;17(8):e1009738. doi: 10.1371/journal.pgen.1009738. eCollection 2021 Aug. PLoS Genet. 2021. PMID: 34411095 Free PMC article.
References
-
- Adams, J. 2004. The development of proteasome inhibitors as anticancer drugs. Cancer Cell 5:417-421. - PubMed
-
- Albino, A. P., M. J. Vidal, N. S. McNutt, C. R. Shea, V. G. Prieto, D. M. Nanus, J. M. Palmer, and N. K. Hayward. 1994. Mutation and expression of the p53 gene in human malignant melanoma. Melanoma Res. 4:35-45. - PubMed
-
- Auvinen, M., A. Paasinen, L. C. Andersson, and E. Holtta. 1992. Ornithine decarboxylase activity is critical for cell transformation. Nature 360:355-358. - PubMed
-
- Baguley, B. C., E. S. Marshall, K. M. Holdaway, G. W. Rewcastle, and W. A. Denny. 1998. Inhibition of growth of primary human tumour cell cultures by a 4-anilinoquinazoline inhibitor of the epidermal growth factor receptor family of tyrosine kinases. Eur. J. Cancer 34:1086-1090. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials