

The molecular chaperone Sse1 and the growth control protein kinase Sch9 collaborate to regulate protein kinase A activity in Saccharomyces cerevisiae
- PMID: 15879503
- PMCID: PMC1451167
- DOI: 10.1534/genetics.105.043109
The molecular chaperone Sse1 and the growth control protein kinase Sch9 collaborate to regulate protein kinase A activity in Saccharomyces cerevisiae
Abstract
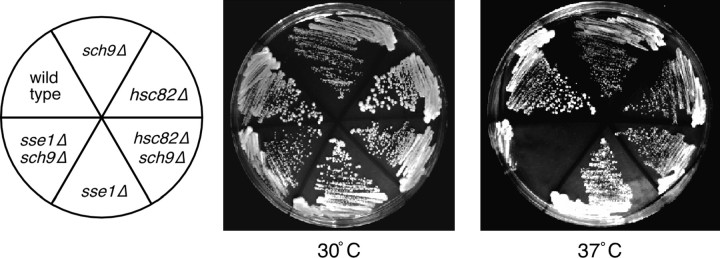
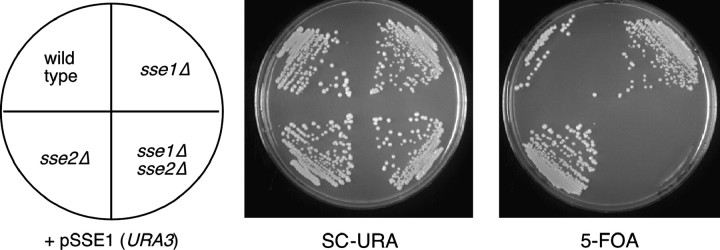
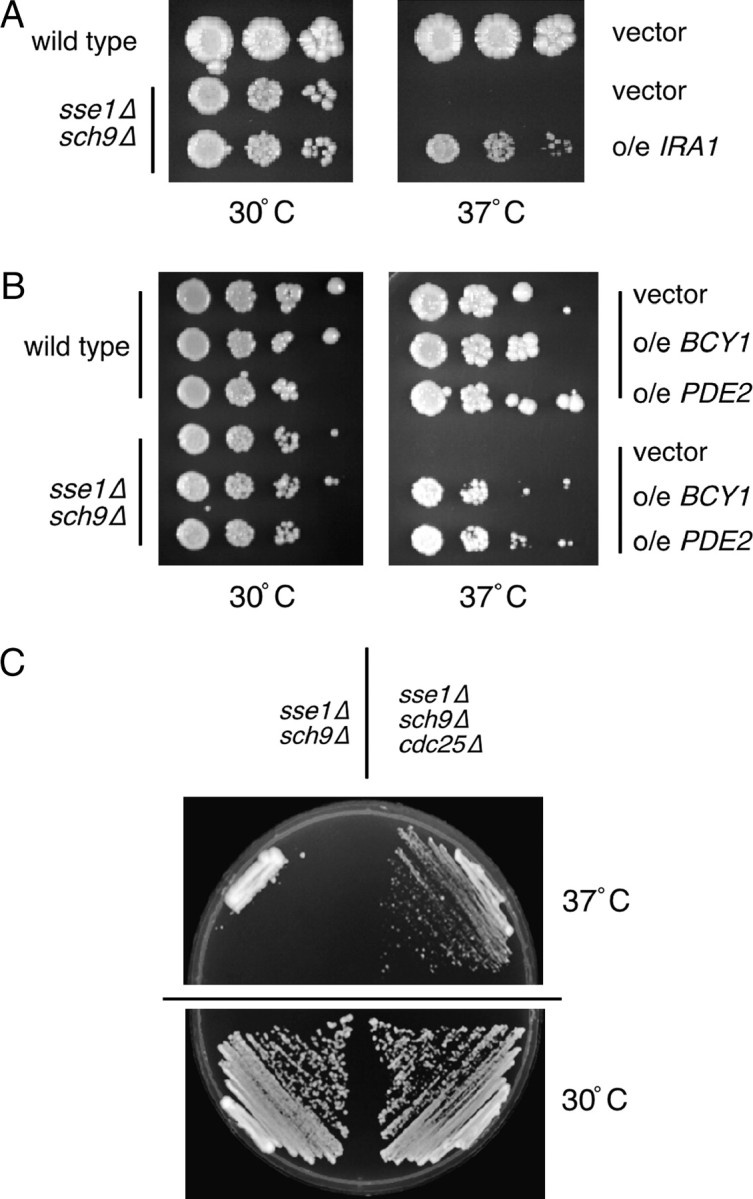
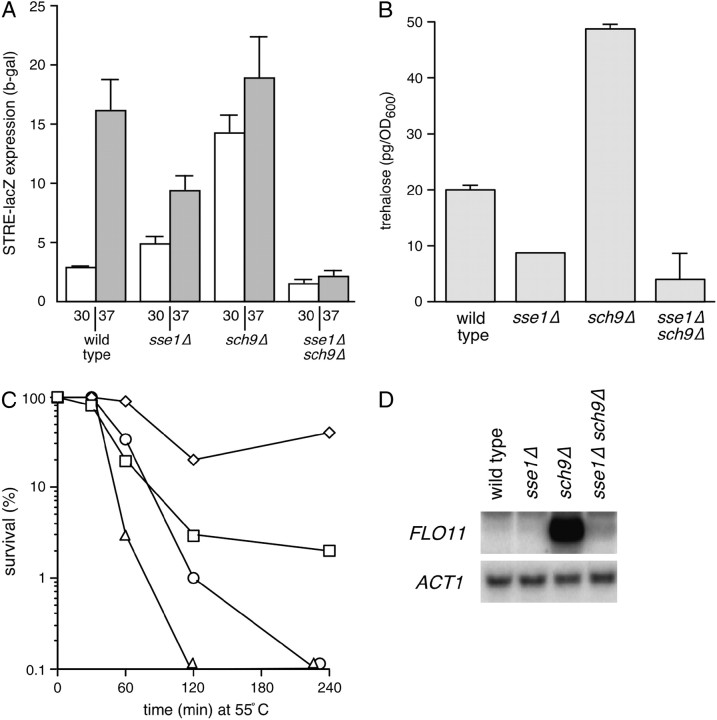
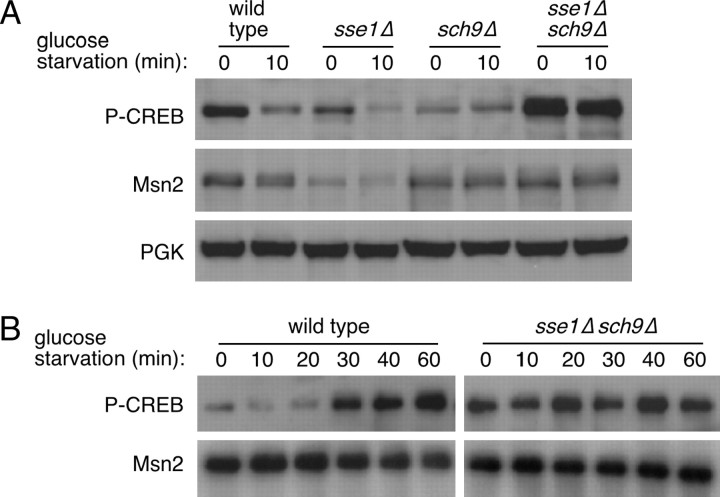
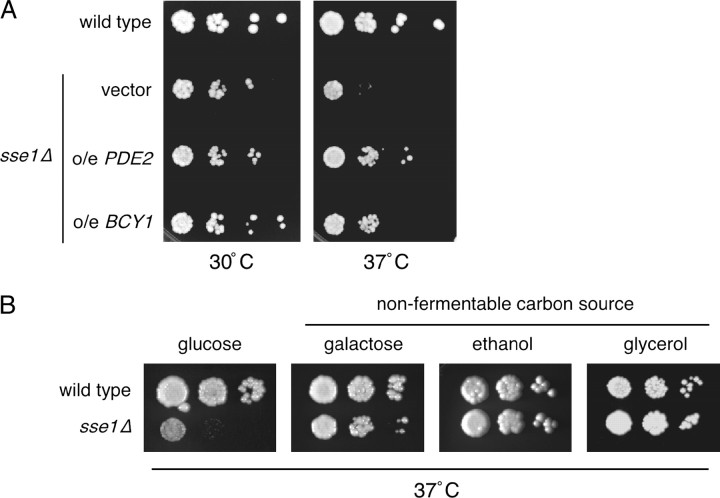
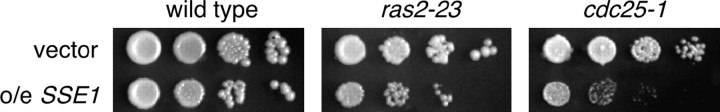
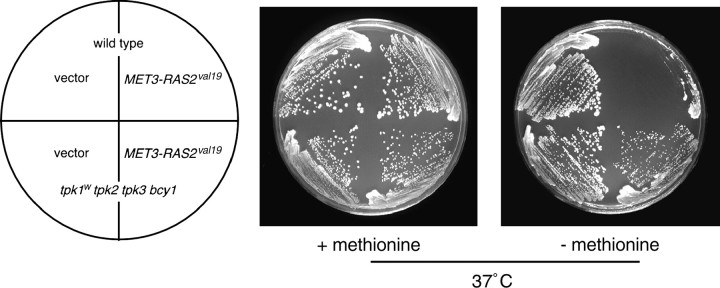
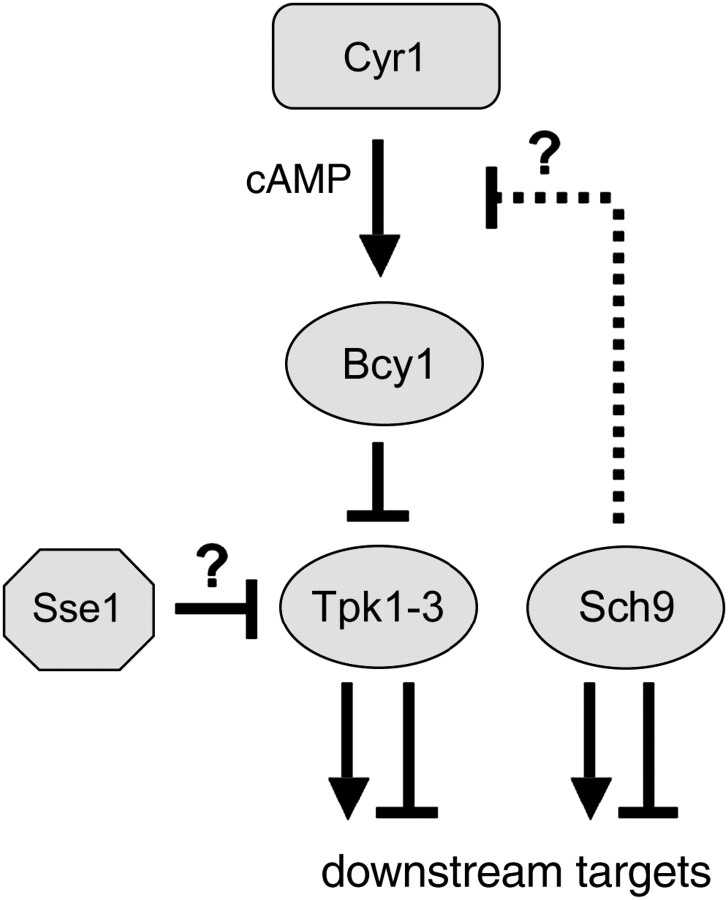
The Sch9 protein kinase regulates Hsp90-dependent signal transduction activity in the budding yeast Saccharomyces cerevisiae. Hsp90 functions in concert with a number of cochaperones, including the Hsp110 homolog Sse1. In this report, we demonstrate a novel synthetic genetic interaction between SSE1 and SCH9. This interaction was observed specifically during growth at elevated temperature and was suppressed by decreased signaling through the protein kinase A (PKA) signal transduction pathway. Correspondingly, sse1Delta sch9Delta cells were shown by both genetic and biochemical approaches to have abnormally high levels of PKA activity and were less sensitive to modulation of PKA by glucose availability. Growth defects of an sse1Delta mutant were corrected by reducing PKA signaling through overexpression of negative regulators or growth on nonoptimal carbon sources. Hyperactivation of the PKA pathway through expression of a constitutive RAS2 allele likewise resulted in temperature-sensitive growth, suggesting that modulation of PKA activity during thermal stress is required for adaptation and viability. Together these results demonstrate that the Sse1 chaperone and the growth control kinase Sch9 independently contribute to regulation of PKA signaling.
Figures
References
-
- Bourbon, H. M., A. Aguilera, A. Z. Ansari, F. J. Asturias, A. J. Berk et al., 2004. A unified nomenclature for protein subunits of mediator complexes linking transcriptional regulators to RNA polymerase II. Mol. Cell 14 553–557. - PubMed
-
- Boy-Marcotte, E., G. Lagniel, M. Perrot, F. Bussereau, A. Boudsocq et al., 1999. The heat shock response in yeast: differential regulations and contributions of the Msn2p/Msn4p and Hsf1p regulons. Mol. Microbiol. 33 274–283. - PubMed
-
- Brodsky, J. L., E. D. Werner, M. E. Dubas, J. L. Goeckeler, K. B. Kruse et al., 1999. The requirement for molecular chaperones during endoplasmic reticulum-associated protein degradation demonstrates that protein export and import are mechanistically distinct. J. Biol. Chem. 274 3453–3460. - PubMed
-
- Broek, D., T. Toda, T. Michaeli, L. Levin, C. Birchmeier et al., 1987. The S. cerevisiae CDC25 gene product regulates the Ras/adenylate cyclase pathway. Cell 48 789–799. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
