

Pontine-wave generator activation-dependent memory processing of avoidance learning involves the dorsal hippocampus in the rat
- PMID: 15880522
- PMCID: PMC1224707
- DOI: 10.1002/jnr.20501
Pontine-wave generator activation-dependent memory processing of avoidance learning involves the dorsal hippocampus in the rat
Abstract
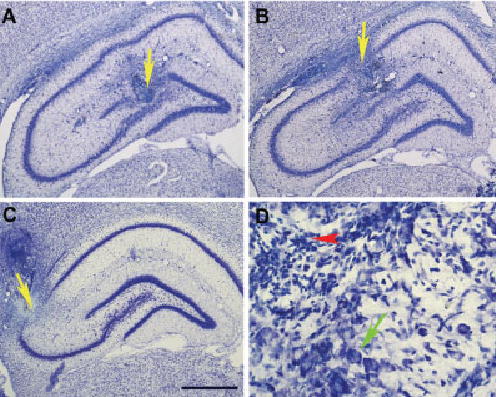
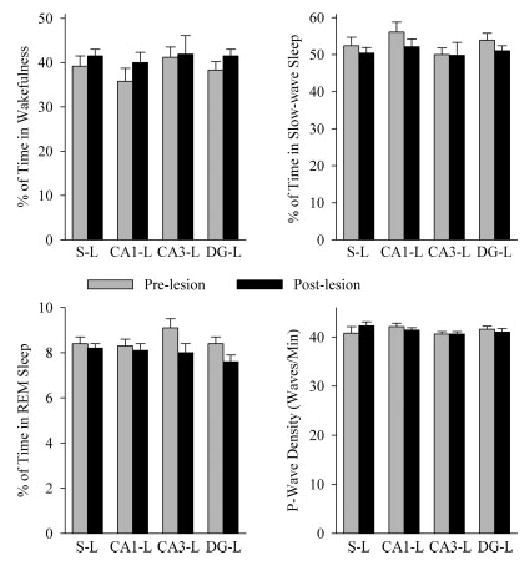
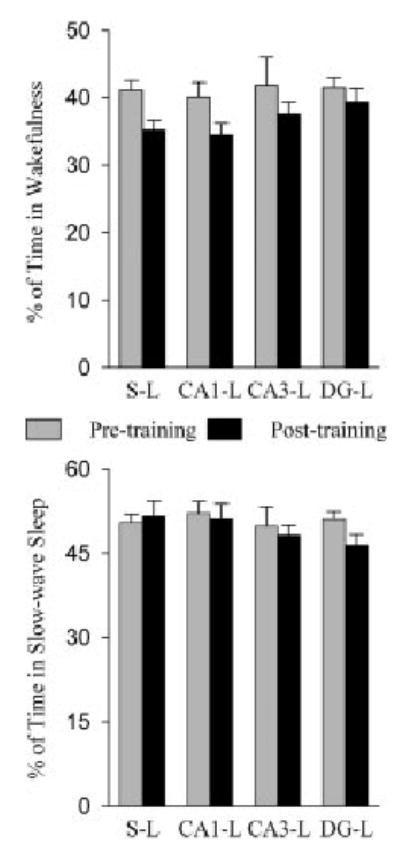
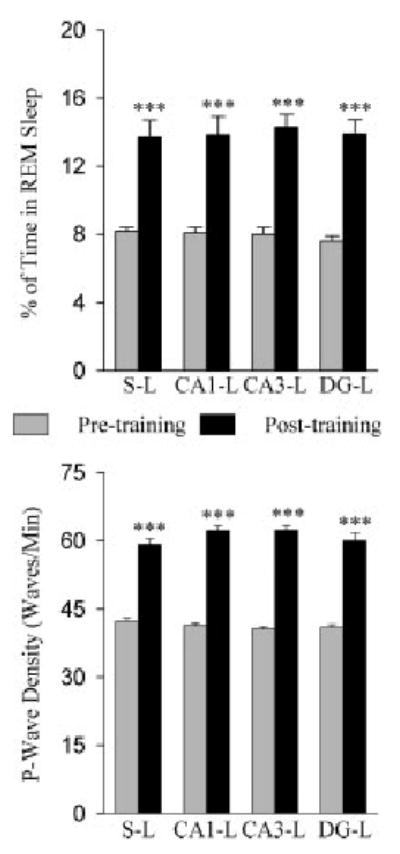
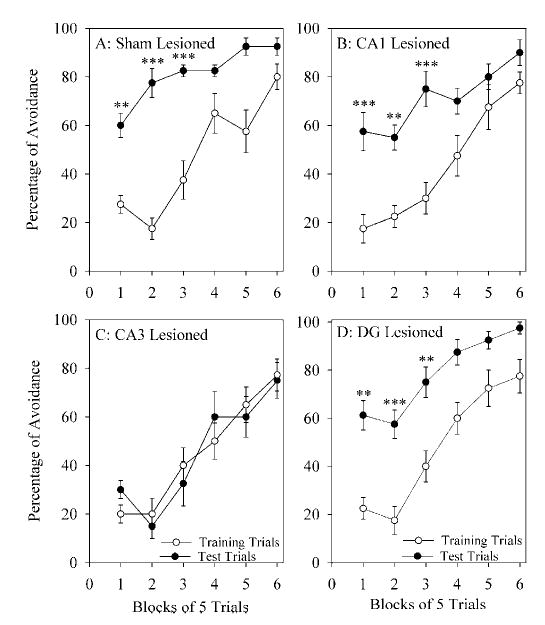
The aim of this study was to test the hypothesis that the dorsal hippocampus plays a critical role in pontine-wave (P-wave) generator activation-dependent memory processing of two-way active avoidance (TWAA) learning. To achieve this objective, rats were given small bilateral lesions in the CA1, dentate gyrus (DG), or CA3 region of the dorsal hippocampus by microinjecting ibotenic acid. After recovery, lesioned and sham-lesioned rats were trained on a TWAA learning paradigm, allowed a 6-hr period of undisturbed sleep, and then were tested on the same TWAA paradigm. It was found that lesions in the CA3 region impaired retention of avoidance learning. Conversely, lesions in the CA1 and DG regions had no effect on TWAA learning retention. None of the groups showed any changes in the baseline sleep-wake cycle or in the acquisition of TWAA learning. All rats showed increased rapid eye movement (REM) sleep and increased REM sleep P-wave density during the subsequent 6-hr recording period. Impaired retention in the CA3 group occurred despite an increase in REM sleep and P-wave density, suggesting that during REM sleep, the P-wave generator interacts with the CA3 region of the dorsal hippocampus to aid in consolidation of TWAA learning. The results of the present study thus demonstrate that P-wave generator activation-dependent consolidation of memory requires an intact CA3 subfield of the dorsal hippocampus. The results also provide evidence that under mnemonic pressure, the dorsal hippocampus may not be involved directly in regulating the sleep-wake cycle.
(c) 2005 Wiley-Liss, Inc.
Figures
References
-
- Aznar S, Rasmussen T, Zimmer J. Impaired learning correlates with size of excitotoxic hippocampal CA3 lesions in adult rats, but shows no amelioration by CA3 transplants. Restor Neurol Neurosci. 1998;13:141–151. - PubMed
-
- Bailey EL, Overstreet DH, Crocker AD. Effects of intrahippocam-pal injections of the cholinergic neurotoxin AF64A on open-field activity and avoidance learning in the rat. Behav Neural Biol. 1986;45:263–274. - PubMed
-
- Bannerman DM, Grubb M, Deacon RM, Yee BK, Feldon J, Rawlins JN. Ventral hippocampal lesions affect anxiety but not spatial learning. Behav Brain Res. 2003;139:197–213. - PubMed
-
- Bannerman DM, Yee BK, Good MA, Heupel MJ, Iversen SD, Rawlins JN. Double dissociation of function within the hippocampus: a comparison of dorsal, ventral, and complete hippocampal cytotoxic lesions. Behav Neurosci. 1999;113:1170–1188. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
