

Autosomal dominant retinitis pigmentosa mutations in inosine 5'-monophosphate dehydrogenase type I disrupt nucleic acid binding
- PMID: 15882147
- PMCID: PMC1184561
- DOI: 10.1042/BJ20042051
Autosomal dominant retinitis pigmentosa mutations in inosine 5'-monophosphate dehydrogenase type I disrupt nucleic acid binding
Abstract
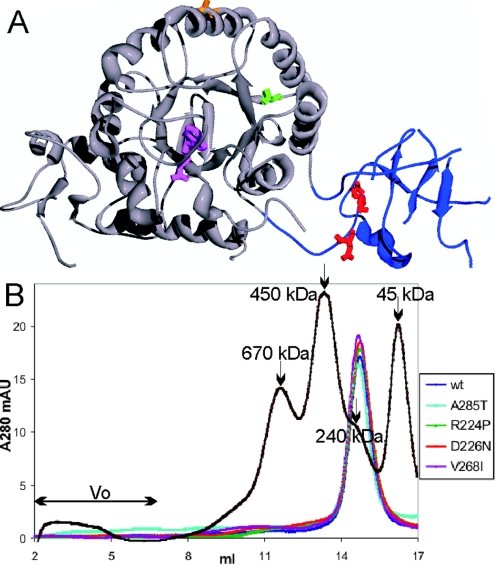
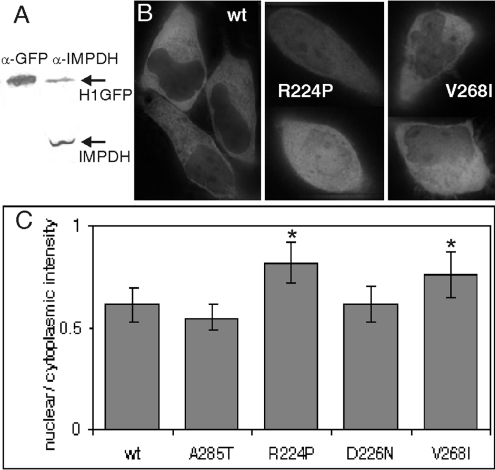
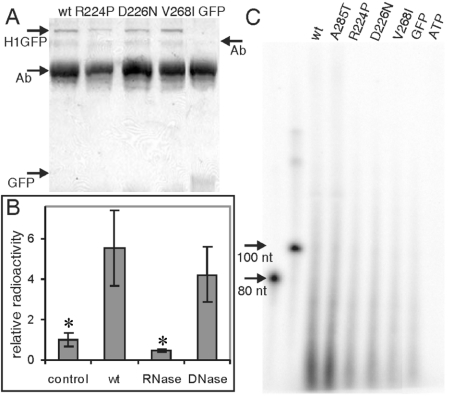
Two mutations of IMPDH1 (inosine 5'-monophosphate dehydrogenase type I), R224P and D226N, have recently been found to cause adRP (autosomal dominant retinitis pigmentosa). IMPDH1 catalyses the rate-limiting step in guanine nucleotide biosynthesis and also binds single-stranded nucleic acids. In the present paper, we report the biochemical characterization of the adRP-linked mutations, R224P and D226N, and a potentially pathogenic mutation, V268I. The adRP-linked mutations have no effect on enzyme activity, protein stability or protein aggregation. These results suggest strongly that the mutations do not affect enzyme activity in vivo and thus do not perturb the guanine nucleotide pool. The R224P mutation changes the distribution of enzyme between the nucleus and cytoplasm. This effect was not observed with the D226N mutation, so the relevance of this observation to disease is unclear. In contrast, both mutations decrease the affinity of nucleic acid binding and both fail to co-immunoprecipitate RNA. These observations suggest that nucleic acid binding provides a functional assay for adRP pathogenicity. The putative adRP-linked mutation V268I also disrupts nucleic acid binding, which suggests that this mutation is indeed pathogenic.
Figures

References
-
- Bunker C. H., Berson E. L., Bromley W. C., Hayes R. P., Roderick T. H. Prevalence of retinitis pigmentosa in Maine. Am. J. Ophthalmol. 1984;97:357–365. - PubMed
-
- Van Soest S., Westerveld A., De Jong P. T., Bleeker-Wagemakers E. M., Bergen A. A. Retinitis pigmentosa: defined from a molecular point of view. Surv. Ophthalmol. 1999;43:321–334. - PubMed
-
- Phelan J. K., Bok D. A brief review of retinitis pigmentosa and the identified retinitis pigmentosa genes. Mol. Vision. 2000;6:116–124. - PubMed
-
- Kennan A., Aherne A., Palfi A., Humphries M., McKee A., Stitt A., Simpson D. A., Demtroder K., Orntoft T., Ayuso C., et al. Identification of an IMPDH1 mutation in autosomal dominant retinitis pigmentosa (RP10) revealed following comparative microarray analysis of transcripts derived from retinas of wild-type and Rho−/− mice. Hum. Mol. Genet. 2002;11:547–558. - PubMed
-
- Bowne S. J., Sullivan L. S., Blanton S. H., Cepko C. L., Blackshaw S., Birch D. G., Hughbanks-Wheaton D., Heckenlively J. R., Daiger S. P. Mutations in the inosine monophosphate dehydrogenase 1 gene (IMPDH1) cause the RP10 form of autosomal dominant retinitis pigmentosa. Hum. Mol. Genet. 2002;11:559–568. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials