

Promotion of importin alpha-mediated nuclear import by the phosphorylation-dependent binding of cargo protein to 14-3-3
- PMID: 15883195
- PMCID: PMC2171942
- DOI: 10.1083/jcb.200411169
Promotion of importin alpha-mediated nuclear import by the phosphorylation-dependent binding of cargo protein to 14-3-3
Abstract
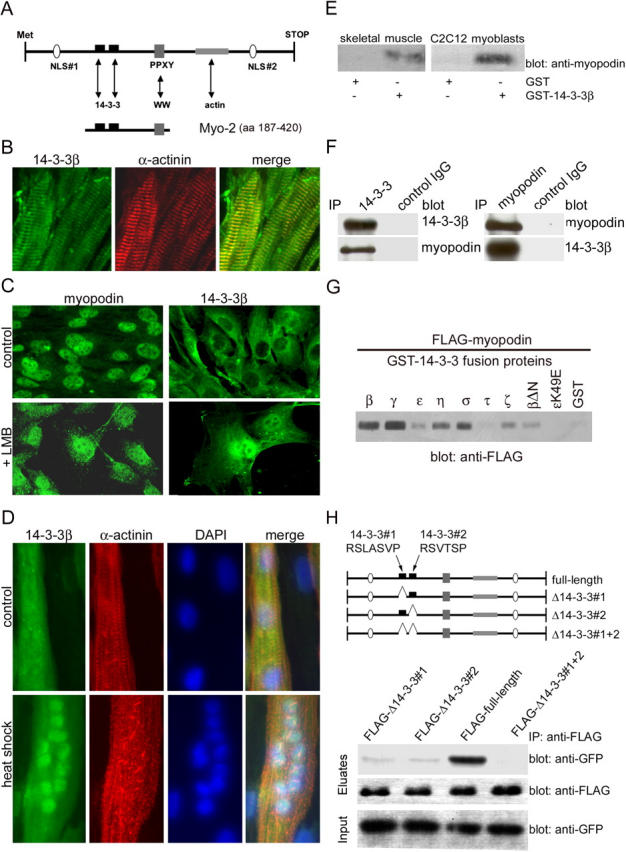
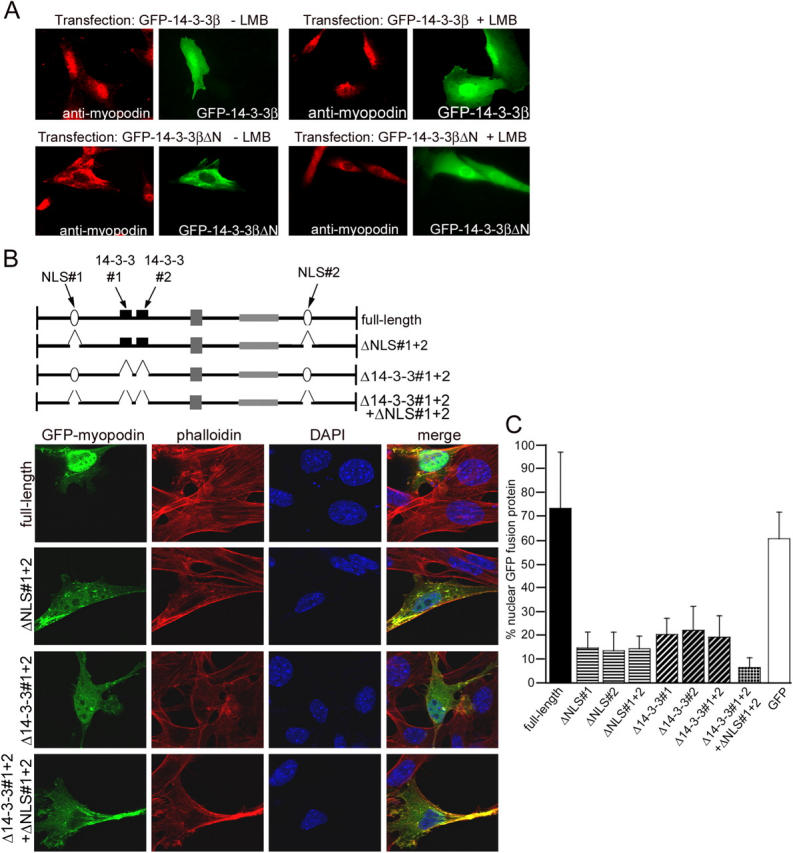
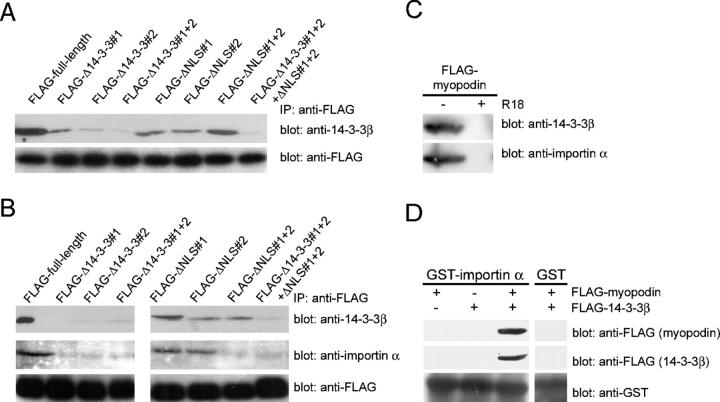
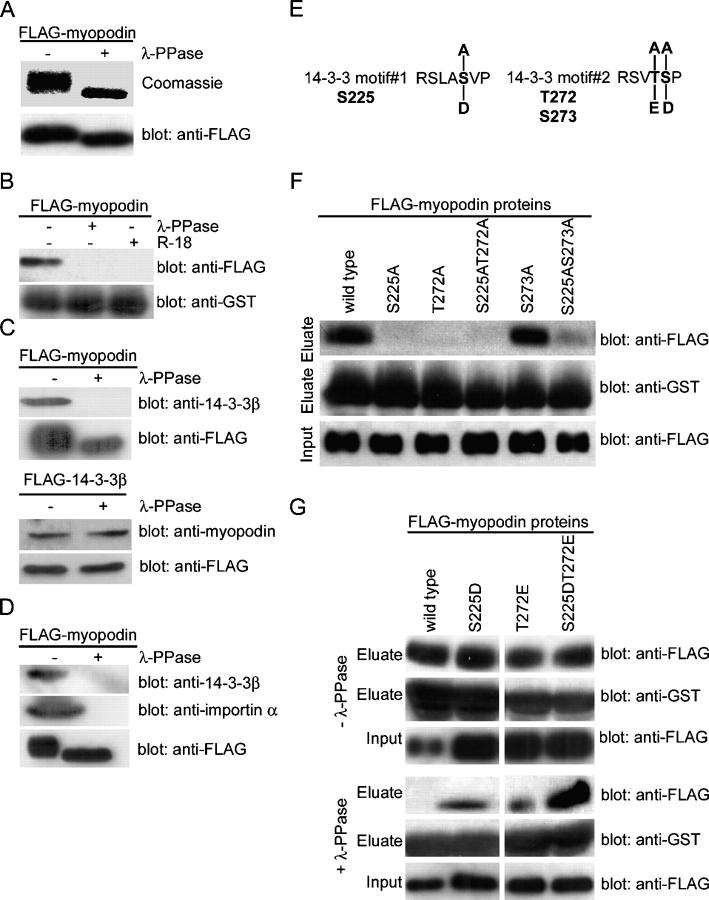
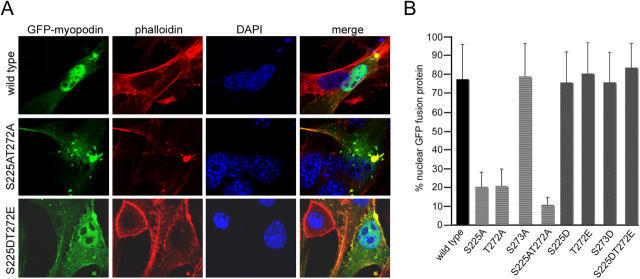
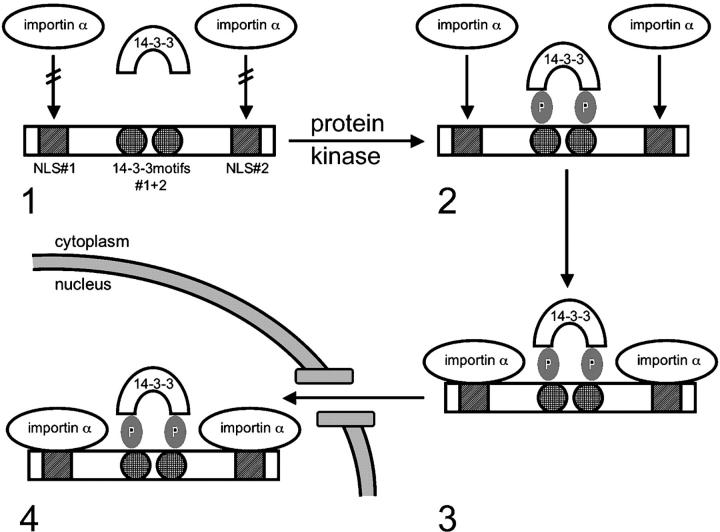
14-3-3 proteins are phosphoserine/threonine-binding proteins that play important roles in many regulatory processes, including intracellular protein targeting. 14-3-3 proteins can anchor target proteins in the cytoplasm and in the nucleus or can mediate their nuclear export. So far, no role for 14-3-3 in mediating nuclear import has been described. There is also mounting evidence that nuclear import is regulated by the phosphorylation of cargo proteins, but the underlying mechanism remains elusive. Myopodin is a dual-compartment, actin-bundling protein that functions as a tumor suppressor in human bladder cancer. In muscle cells, myopodin redistributes between the nucleus and the cytoplasm in a differentiation-dependent and stress-induced fashion. We show that importin alpha binding and the subsequent nuclear import of myopodin are regulated by the serine/threonine phosphorylation-dependent binding of myopodin to 14-3-3. These results establish a novel paradigm for the promotion of nuclear import by 14-3-3 binding. They provide a molecular explanation for the phosphorylation-dependent nuclear import of nuclear localization signal-containing cargo proteins.
Figures
Similar articles
-
Protein kinase A, Ca2+/calmodulin-dependent kinase II, and calcineurin regulate the intracellular trafficking of myopodin between the Z-disc and the nucleus of cardiac myocytes.Mol Cell Biol. 2007 Dec;27(23):8215-27. doi: 10.1128/MCB.00950-07. Epub 2007 Oct 8. Mol Cell Biol. 2007. PMID: 17923693 Free PMC article.
-
Differentiation- and stress-dependent nuclear cytoplasmic redistribution of myopodin, a novel actin-bundling protein.J Cell Biol. 2001 Oct 29;155(3):393-404. doi: 10.1083/jcb.200012039. Epub 2001 Oct 22. J Cell Biol. 2001. PMID: 11673475 Free PMC article.
-
Nuclear Respiratory Factor 2β (NRF-2β) recruits NRF-2α to the nucleus by binding to importin-α:β via an unusual monopartite-type nuclear localization signal.J Mol Biol. 2013 Sep 23;425(18):3536-48. doi: 10.1016/j.jmb.2013.07.007. Epub 2013 Jul 13. J Mol Biol. 2013. PMID: 23856623
-
Nuclear import and export of proteins in plants: a tool for the regulation of signalling.Planta. 2001 Aug;213(4):499-517. doi: 10.1007/s004250100621. Planta. 2001. PMID: 11556782 Review. No abstract available.
-
[Functional Dynamics of Importin α family].Seikagaku. 2015 Feb;87(1):16-21. Seikagaku. 2015. PMID: 26571550 Review. Japanese. No abstract available.
Cited by
-
Rescue of tropomyosin deficiency in Drosophila and human cancer cells by synaptopodin reveals a role of tropomyosin α in RhoA stabilization.EMBO J. 2012 Feb 15;31(4):1028-40. doi: 10.1038/emboj.2011.464. Epub 2011 Dec 13. EMBO J. 2012. PMID: 22157816 Free PMC article.
-
The sarcomeric Z-disc: a nodal point in signalling and disease.J Mol Med (Berl). 2006 Jun;84(6):446-68. doi: 10.1007/s00109-005-0033-1. Epub 2006 Jan 17. J Mol Med (Berl). 2006. PMID: 16416311 Review.
-
Nuclear relocation of the nephrin and CD2AP-binding protein dendrin promotes apoptosis of podocytes.Proc Natl Acad Sci U S A. 2007 Jun 12;104(24):10134-9. doi: 10.1073/pnas.0700917104. Epub 2007 May 30. Proc Natl Acad Sci U S A. 2007. PMID: 17537921 Free PMC article.
-
hnRNPA1 couples nuclear export and translation of specific mRNAs downstream of FGF-2/S6K2 signalling.Nucleic Acids Res. 2014 Nov 10;42(20):12483-97. doi: 10.1093/nar/gku953. Epub 2014 Oct 16. Nucleic Acids Res. 2014. PMID: 25324306 Free PMC article.
-
14-3-3zeta escorts CCTalpha for calcium-activated nuclear import in lung epithelia.FASEB J. 2010 Apr;24(4):1271-83. doi: 10.1096/fj.09-136044. Epub 2009 Dec 9. FASEB J. 2010. PMID: 20007511 Free PMC article.
References
-
- Alam, R., N. Hachiya, M. Sakaguchi, S. Kawabata, S. Iwanaga, M. Kitajima, K. Mihara, and T. Omura. 1994. cDNA cloning and characterization of mitochondrial import stimulation factor (MSF) purified from rat liver cytosol. J. Biochem. (Tokyo). 116:416–425. - PubMed
-
- Bihn, E.A., A.L. Paul, S.W. Wang, G.W. Erdos, and R.J. Ferl. 1997. Localization of 14-3-3 proteins in the nuclei of arabidopsis and maize. Plant J. 12:1439–1445. - PubMed
-
- Briggs, L.J., D. Stein, J. Goltz, V.C. Corrigan, A. Efthymiadis, S. Hubner, and D.A. Jans. 1998. The cAMP-dependent protein kinase site (Ser312) enhances dorsal nuclear import through facilitating nuclear localization sequence/importin interaction. J. Biol. Chem. 273:22745–22752. - PubMed
-
- Cahill, C.M., G. Tzivion, N. Nasrin, S. Ogg, J. Dore, G. Ruvkun, and M. Alexander-Bridges. 2001. Phosphatidylinositol 3-kinase signaling inhibits DAF-16 DNA binding and function via 14-3-3-dependent and 14-3-3-independent pathways. J. Biol. Chem. 276:13402–13410. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
