

Clathrin heavy and light chain isoforms originated by independent mechanisms of gene duplication during chordate evolution
- PMID: 15883369
- PMCID: PMC1091751
- DOI: 10.1073/pnas.0502058102
Clathrin heavy and light chain isoforms originated by independent mechanisms of gene duplication during chordate evolution
Abstract
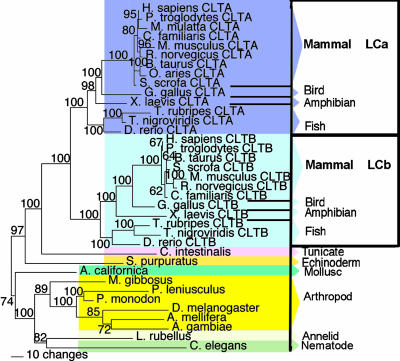
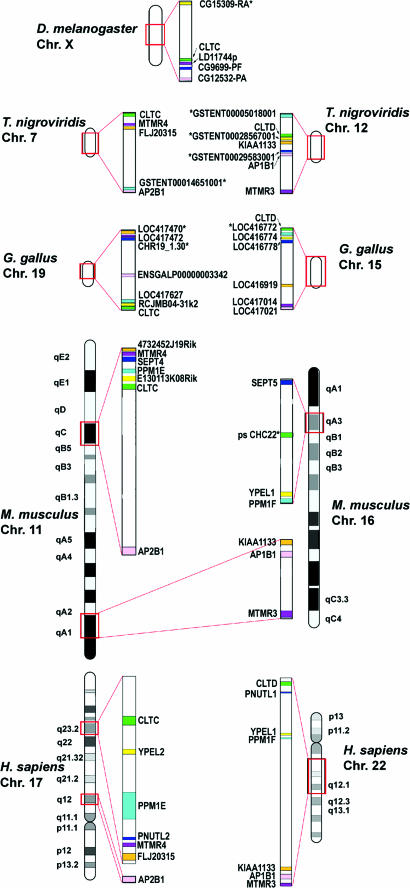
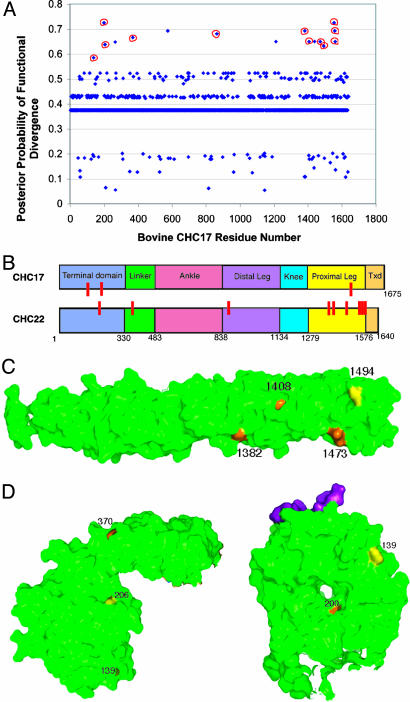
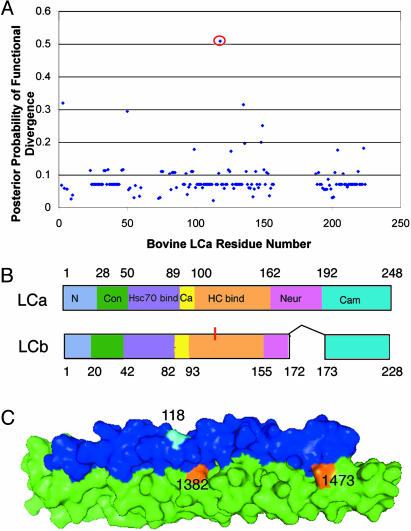
In humans, there are two isoforms each of clathrin heavy chain (CHC17 and CHC22) and light chain (LCa and LCb) subunits, all encoded by separate genes. CHC17 forms the ubiquitous clathrin-coated vesicles that mediate membrane traffic. CHC22 is implicated in specialized membrane organization in skeletal muscle. CHC17 is bound and regulated by LCa and LCb, whereas CHC22 does not functionally interact with either light chain. The imbalanced interactions between clathrin subunit isoforms suggest a distinct evolutionary history for each isoform pair. Phylogenetic and sequence analysis placed both heavy and light chain gene duplications during chordate evolution, 510-600 million years ago. Genes encoding CHC22 orthologues were found in several vertebrate species, with only a pseudogene present in mice. Multiple paralogons surrounding the CHC genes (CLTC and CLTD) were identified, evidence that genomic or large-scale gene duplication produced the two CHC isoforms. In contrast, clathrin light chain genes (CLTA and CLTB) apparently arose by localized duplication, within 1-11 million years of CHC gene duplication. Analysis of sequence divergence patterns suggested that structural features of the CHCs were maintained after gene duplication, but new interactions with regulatory proteins evolved for the CHC22 isoform. Thus, independent mechanisms of gene duplication expanded clathrin functions, concomitant with development of neuromuscular sophistication in chordates.
Figures

References
-
- Ferrier, D. E. & Minguillon, C. (2003) Int. J. Dev. Biol. 47, 605-611. - PubMed
-
- Yu, C. Y. (1998) Exp. Clin. Immunogenet. 15, 213-230. - PubMed
-
- Schledzewski, K., Brinkmann, H. & Mendel, R. R. (1999) J. Mol. Evol. 48, 770-778. - PubMed
-
- Dodge, G. R., Kovalszky, I., McBride, O. W., Yi, H. J., Chu, M., Saitta, B., Stokes, D. G. & Ioozo, R. V. (1991) Genomics 11, 174-178. - PubMed
-
- Sirotkin, H., Morrow, B., DasGupta, R., Goldberg, R., Patanjali, S. R., Shi, G., Cannizzaro, L., Shprintzen, R., Weissman, S. M. & Kucherlapati, R. (1996) Hum. Mol. Genet. 5, 617-624. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
