

The negative regulatory function of the lymphocyte-activation gene-3 co-receptor (CD223) on human T cells
- PMID: 15885122
- PMCID: PMC1782137
- DOI: 10.1111/j.1365-2567.2005.02145.x
The negative regulatory function of the lymphocyte-activation gene-3 co-receptor (CD223) on human T cells
Abstract
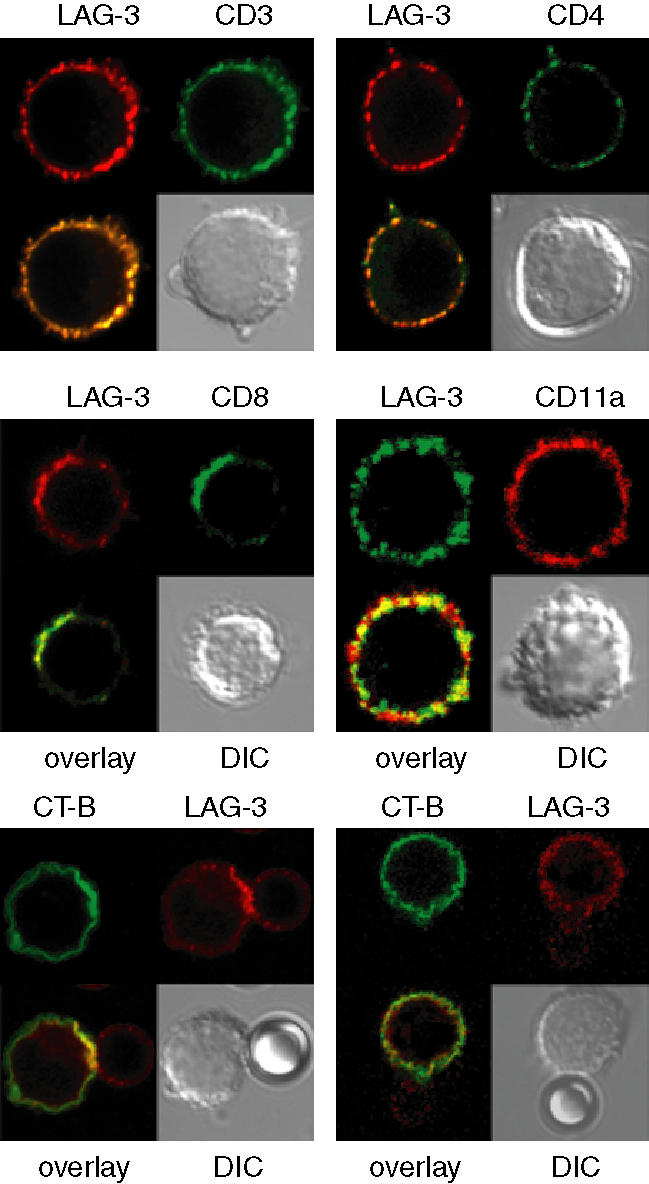
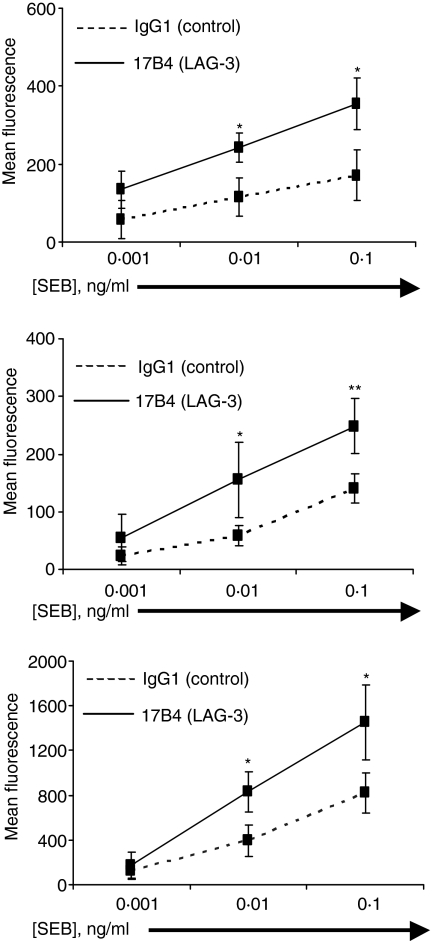
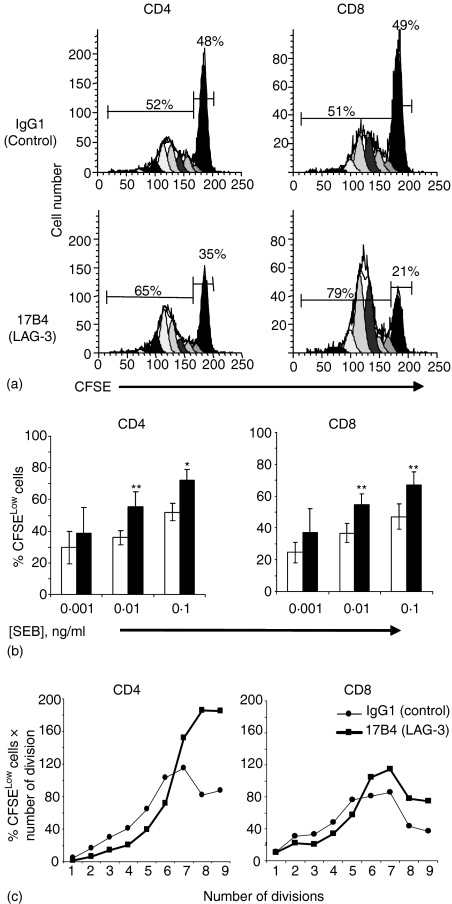
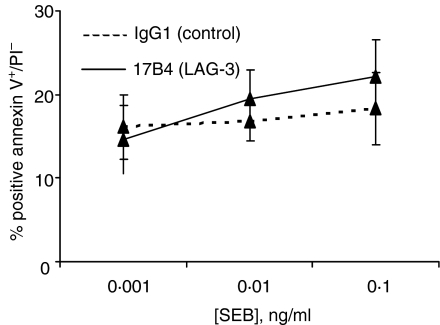
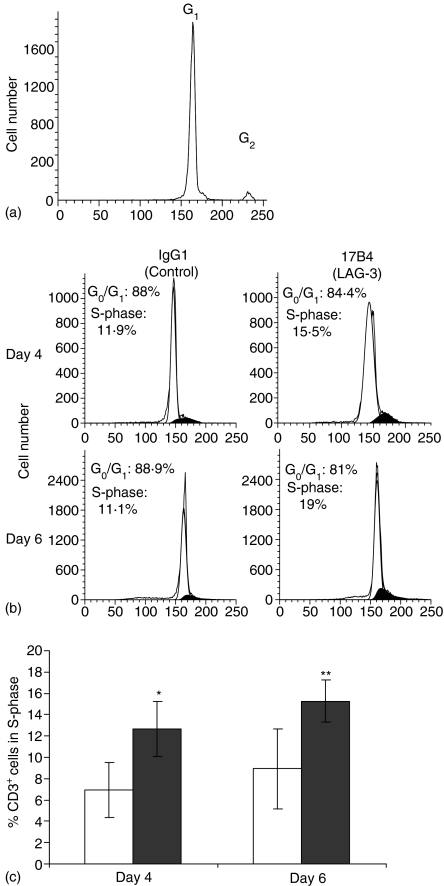
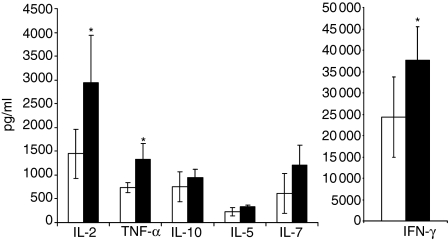
Accumulating evidence indicates that the CD4 homologue lymphocyte activation gene-3 (LAG-3) plays a down-regulatory role on T-cell responses. However, the role of LAG-3/major histocompatibility complex (MHC) class II interactions on primary human T-cell responses, as well as the mechanism by which down-regulation occurs, are not clear. Here, we show that LAG-3 colocalized with CD3, CD4 or CD8 in areas of cholesterol-rich raft aggregation during this primary response, as well as in the clustered raft region formed between T cells and antibody-coated beads. Addition of a blocking LAG-3-specific monoclonal antibody to both CD4 and CD8 primary resting T cells activated under conditions of antigen-presenting cell-driven stimulation and low antigen concentrations augments CD69 activation antigen expression, T-cell expansion and T helper 1 (Th1, but not Th2) cytokine production. Blocking LAG-3/MHC class II interactions leads to an increase in the number of cells entering division at these low concentrations of antigen and to more rounds of divisions with an accumulation of cells in the S-phase of the cell cycle. These results indicate that LAG-3 signalling inhibits early events in primary activation of human CD4 and CD8 T cells and further support a role for LAG-3 signalling in regulating the expansion of activated effector or memory T cells, either directly or indirectly through Treg suppressor activity.
Figures
References
-
- Viola A, Schroeder S, Sakakibara Y, Lanzavecchia A. T lymphocyte costimulation mediated by reorganization of membrane microdomains. Science. 1999;283:680–2. - PubMed
-
- Xavier R, Brennan T, Li Q, McCormack C, Seed B. Membrane compartmentation is required for efficient T cell activation. Immunity. 1998;8:723–32. - PubMed
-
- Bi K, Tanaka Y, Coudronniere N, Sugie K, Hong S, van Stipdonk MJ, Altman A. Antigen-induced translocation of PKC-theta to membrane rafts is required for T cell activation. Nat Immunol. 2001;2:556–63. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
