

Specific features of neuronal size and shape are regulated by tropomyosin isoforms
- PMID: 15888546
- PMCID: PMC1165423
- DOI: 10.1091/mbc.e04-10-0951
Specific features of neuronal size and shape are regulated by tropomyosin isoforms
Abstract
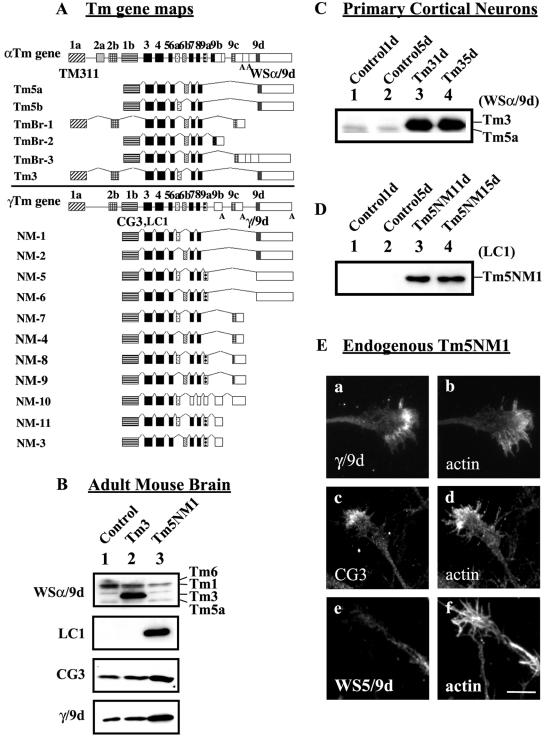
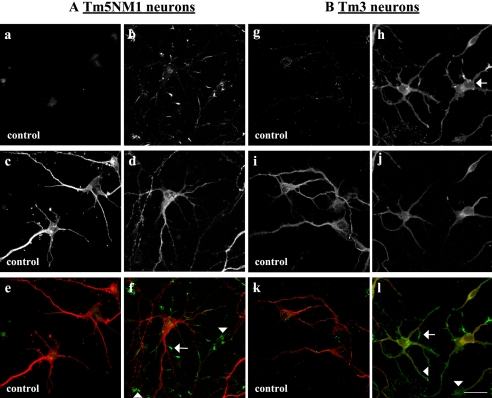
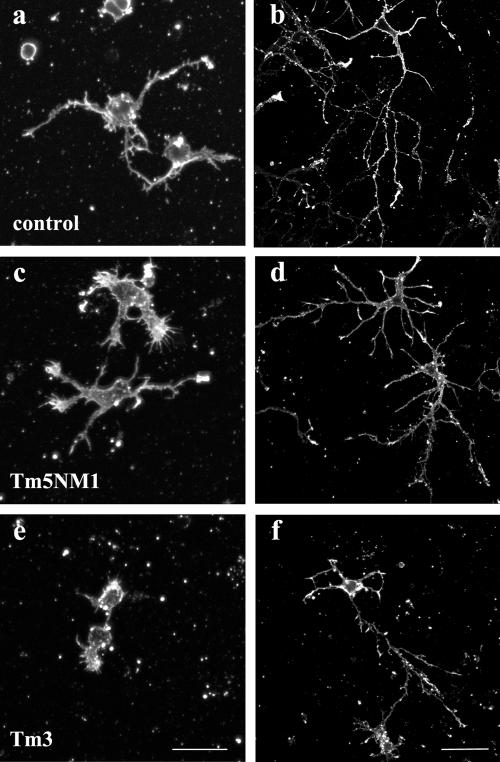
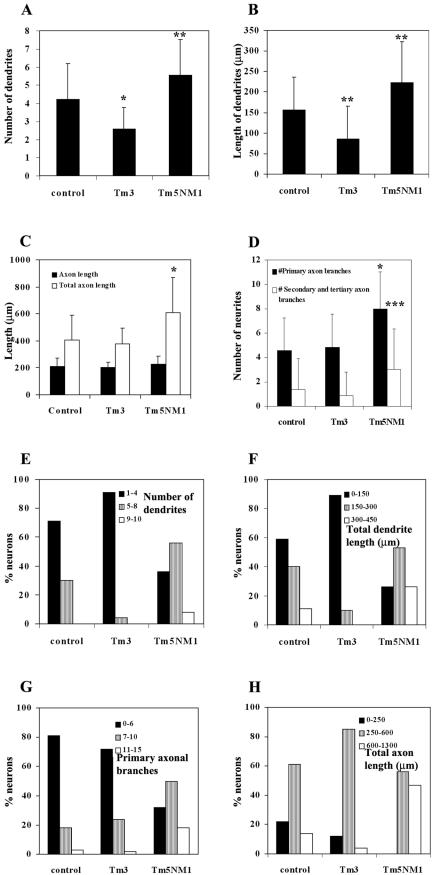
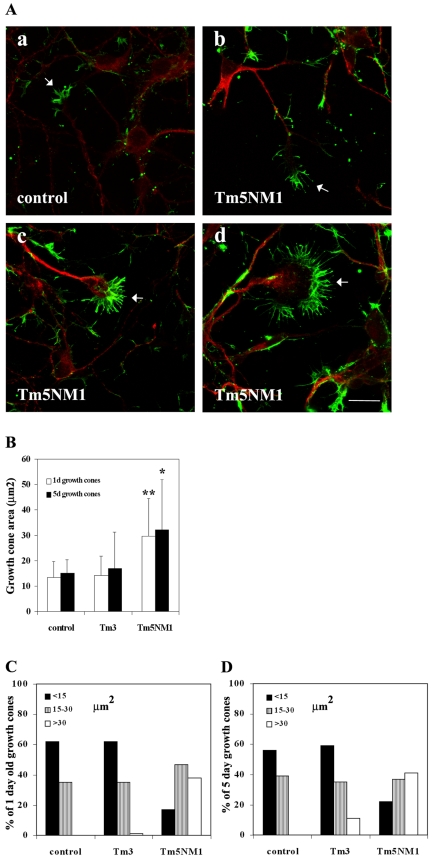
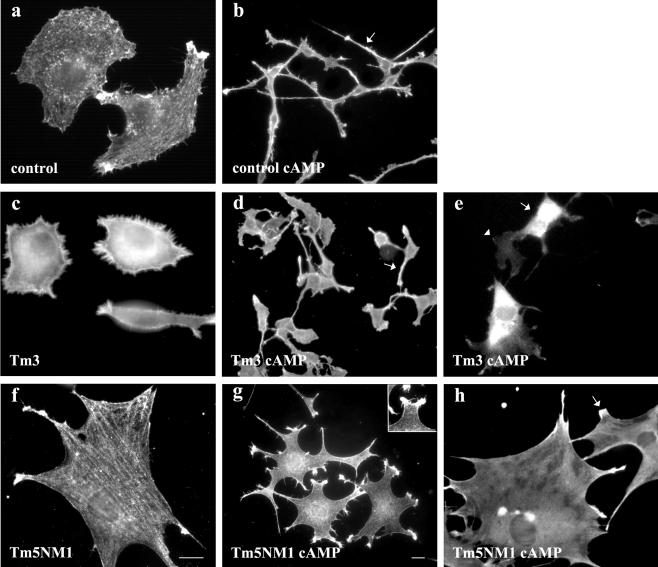
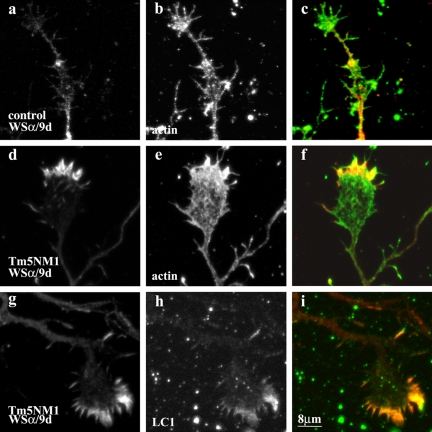
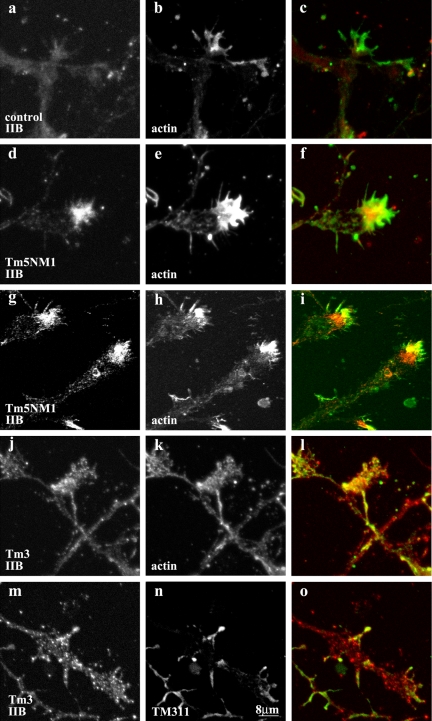
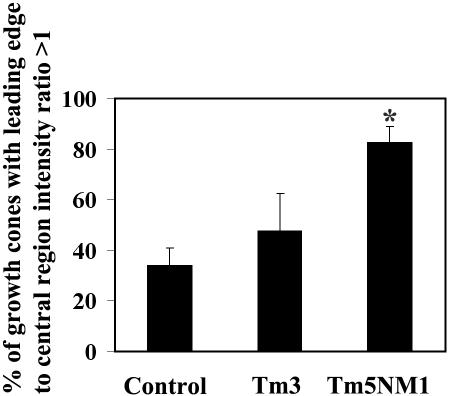
Spatially distinct populations of microfilaments, characterized by different tropomyosin (Tm) isoforms, are present within a neuron. To investigate the impact of altered tropomyosin isoform expression on neuronal morphogenesis, embryonic cortical neurons from transgenic mice expressing the isoforms Tm3 and Tm5NM1, under the control of the beta-actin promoter, were cultured in vitro. Exogenously expressed Tm isoforms sorted to different subcellular compartments with Tm5NM1 enriched in filopodia and growth cones, whereas the Tm3 was more broadly localized. The Tm5NM1 neurons displayed significantly enlarged growth cones accompanied by an increase in the number of dendrites and axonal branching. In contrast, Tm3 neurons displayed inhibition of neurite outgrowth. Recruitment of Tm5a and myosin IIB was observed in the peripheral region of a significant number of Tm5NM1 growth cones. We propose that enrichment of myosin IIB increases filament stability, leading to the enlarged growth cones. Our observations support a role for different tropomyosin isoforms in regulating interactions with myosin and thereby regulating morphology in specific intracellular compartments.
Figures
References
-
- Bernstein, B. W., and Bamburg, J. R. (1982). Tropomyosin binding to F-actin protects the F-actin from disassembly by brain actin-depolymerizing factor (ADF). Cell Motil. 2, 1–8. - PubMed
-
- Cooley, B. C., and Bergtrom, G. (2001). Multiple combinations of alternatively spliced exons in rat tropomyosin-α gene mRNA: evidence for 20 new isoforms in adult tissues and cultured cells. Arch. Biochem. Biophys. 390, 71–77. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
