

Prostaglandin D2 induces nuclear import of the sex-determining factor SOX9 via its cAMP-PKA phosphorylation
- PMID: 15889150
- PMCID: PMC1142593
- DOI: 10.1038/sj.emboj.7600660
Prostaglandin D2 induces nuclear import of the sex-determining factor SOX9 via its cAMP-PKA phosphorylation
Abstract
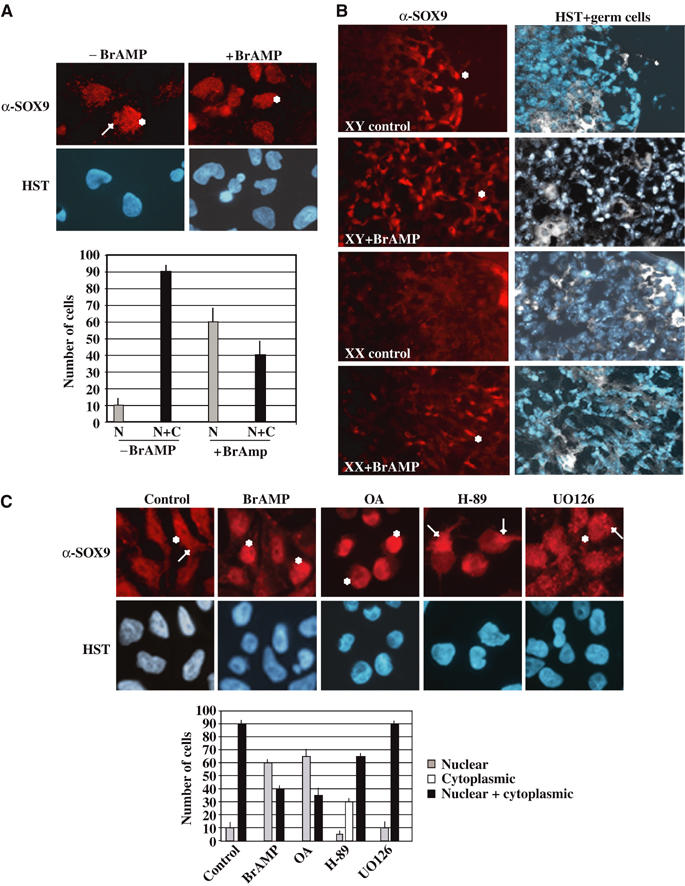
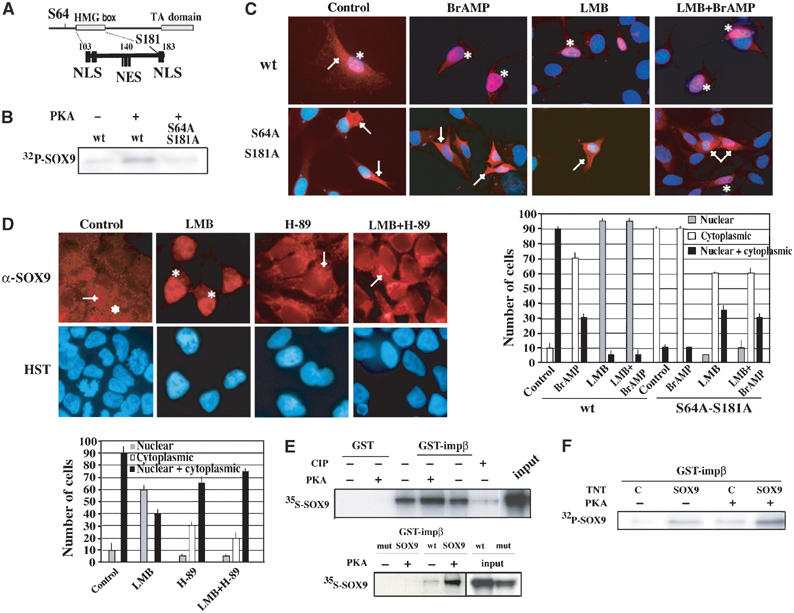
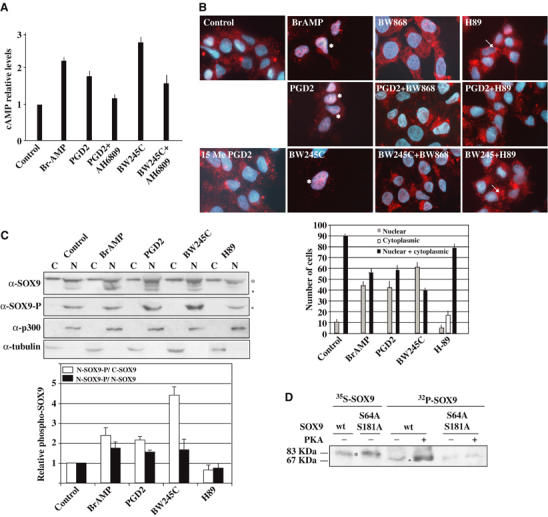
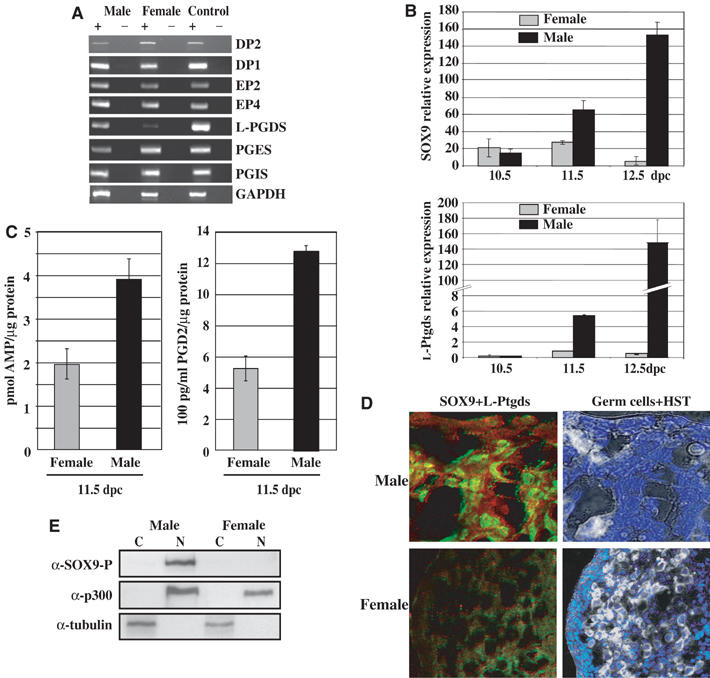
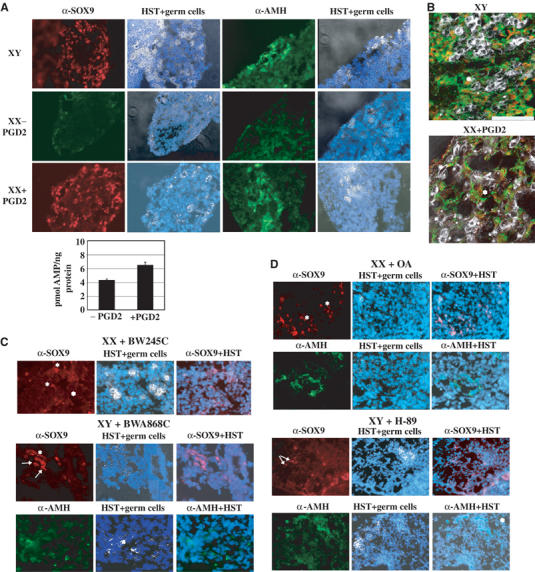
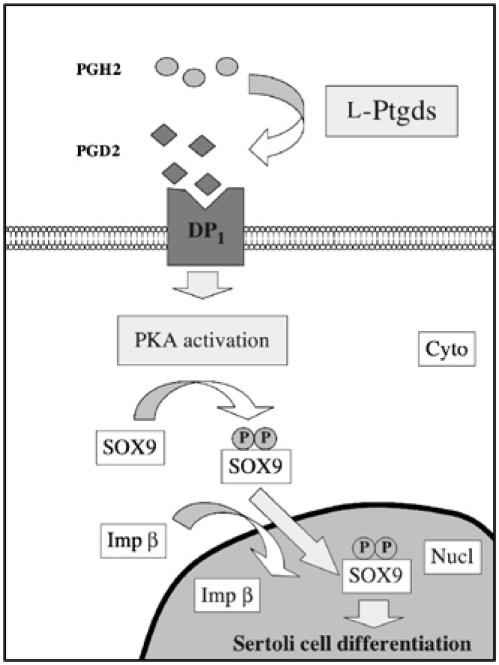
During mammalian gonadal development, nuclear import/export of the transcription factor SOX9 is a critical step of the Sry-initiated testis-determining cascade. In this study, we identify a molecular mechanism contributing to the SOX9 nuclear translocation in NT2/D1 cells, which is mediated by the prostaglandin D2 (PGD2) signalling pathway via stimulation of its adenylcyclase-coupled DP1 receptor. We find that activation of cAMP-dependent protein kinase A (PKA) induces phosphorylation of SOX9 on its two S64 and S181 PKA sites, and its nuclear localization by enhancing SOX9 binding to the nucleocytoplasmic transport protein importin beta. Moreover, in embryonic gonads, we detect a male-specific prostaglandin D synthase expression and an active PGD2 signal at the time and place of SOX9 expression. We thus propose a new step in the sex-determining cascade where PGD2 acts as an autocrine factor inducing SOX9 nuclear translocation and subsequent Sertoli cell differentiation.
Figures
References
-
- Adams IR, McLaren A (2002) Sexually dimorphic development of mouse primordial germ cells: switching from oogenesis to spermatogenesis. Development 129: 1155–1164 - PubMed
-
- Arango NA, Lovell-Badge R, Behringer RR (1999) Targeted mutagenesis of the endogenous mouse Mis gene promoter: in vivo definition of genetic pathways of vertebrate sexual development. Cell 99: 409–419 - PubMed
-
- Argentaro A, Sim H, Kelly S, Preiss S, Clayton A, Jans DA, Harley VR (2003) A SOX9 defect of calmodulin-dependent nuclear import in campomelic dysplasia/autosomal sex reversal. J Biol Chem 278: 33839–33847 - PubMed
-
- Boie Y, Sawyer N, Slipetz DM, Metters KM, Abramovitz M (1995) Molecular cloning and characterization of the human prostanoid DP receptor. J Biol Chem 270: 18910–18916 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
