

Cellulose biosynthesis: current views and evolving concepts
- PMID: 15894551
- PMCID: PMC4246814
- DOI: 10.1093/aob/mci155
Cellulose biosynthesis: current views and evolving concepts
Abstract
Aims: To outline the current state of knowledge and discuss the evolution of various viewpoints put forth to explain the mechanism of cellulose biosynthesis. *
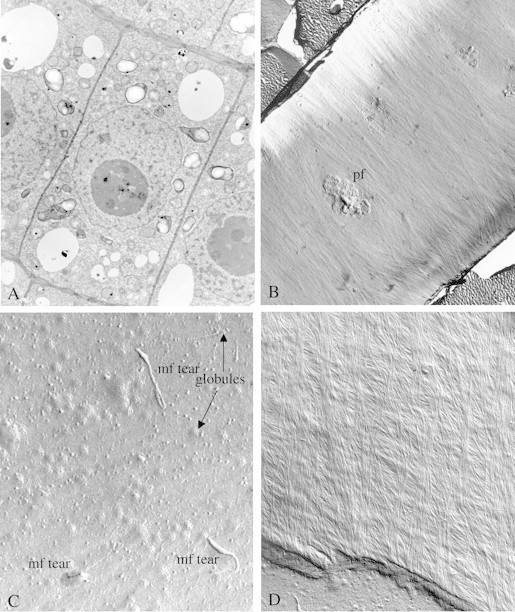
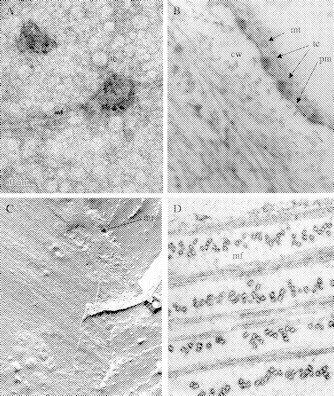
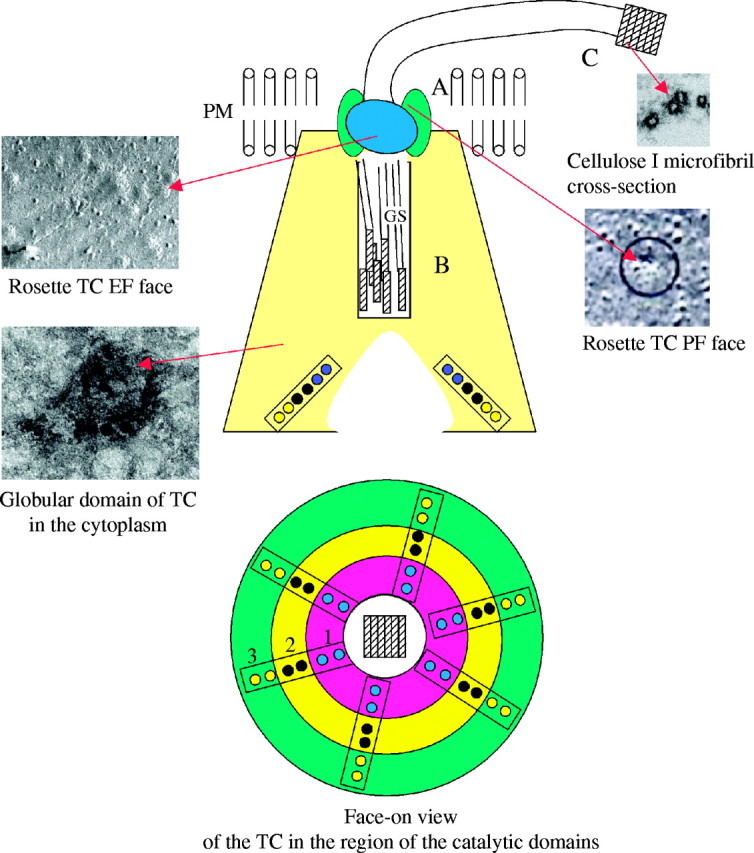
Scope: Understanding the mechanism of cellulose biosynthesis is one of the major challenges in plant biology. The simplicity in the chemical structure of cellulose belies the complexities that are associated with the synthesis and assembly of this polysaccharide. Assembly of cellulose microfibrils in most organisms is visualized as a multi-step process involving a number of proteins with the key protein being the cellulose synthase catalytic sub-unit. Although genes encoding this protein have been identified in almost all cellulose synthesizing organisms, it has been a challenge in general, and more specifically in vascular plants, to demonstrate cellulose synthase activity in vitro. The assembly of glucan chains into cellulose microfibrils of specific dimensions, viewed as a spontaneous process, necessitates the assembly of synthesizing sites unique to most groups of organisms. The steps of polymerization (requiring the specific arrangement and activity of the cellulose synthase catalytic sub-units) and crystallization (directed self-assembly of glucan chains) are certainly interlinked in the formation of cellulose microfibrils. Mutants affected in cellulose biosynthesis have been identified in vascular plants. Studies on these mutants and herbicide-treated plants suggest an interesting link between the steps of polymerization and crystallization during cellulose biosynthesis. *
Conclusions: With the identification of a large number of genes encoding cellulose synthases and cellulose synthase-like proteins in vascular plants and the supposed role of a number of other proteins in cellulose biosynthesis, a complete understanding of this process will necessitate a wider variety of research tools and approaches than was thought to be required a few years back.
Figures
References
-
- Akashi T, Shibaoka. 1991. Involvement of transmembrane proteins in the association of cortical microtubules with the plasma membrane in tobacco BY-2 cells. Journal of Cell Science 98: 169–174.
-
- Aloni Y, Delmer DP, Benziman M. 1982. Achievement of high rates of in vitro synthesis of 1,4-β-glucan: activation by cooperative interaction of the Acetobacter xylinum enzyme system with GTP, polyethylene glycol, and a protein factor. Proceedings of the National Academy of Sciences of the USA 79: 6448–6452. - PMC - PubMed
-
- Appenzeller L, Doblin M, Barreiro R, Wang H, Niu X, Kollipara K, et al. 2004. Cellulose synthesis in maize: isolation and expression analysis of the cellulose synthase (CesA) gene family. Cellulose 11: 287–299.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
