

Arabidopsis thickvein mutation affects vein thickness and organ vascularization, and resides in a provascular cell-specific spermine synthase involved in vein definition and in polar auxin transport
- PMID: 15894745
- PMCID: PMC1150395
- DOI: 10.1104/pp.104.055756
Arabidopsis thickvein mutation affects vein thickness and organ vascularization, and resides in a provascular cell-specific spermine synthase involved in vein definition and in polar auxin transport
Abstract
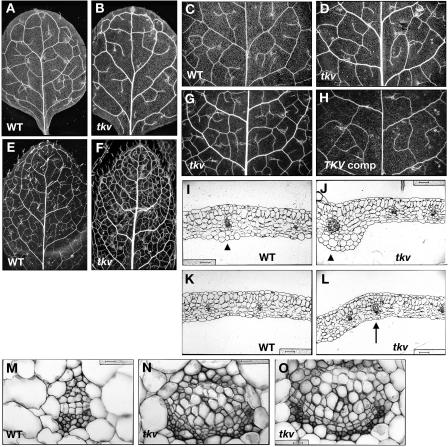
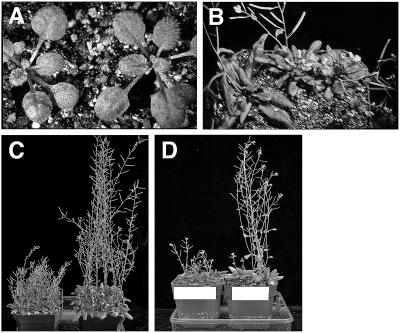
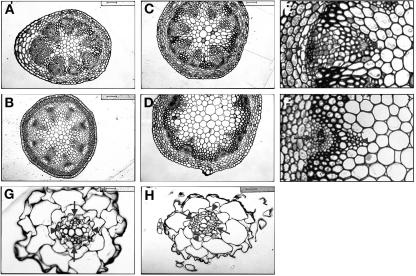
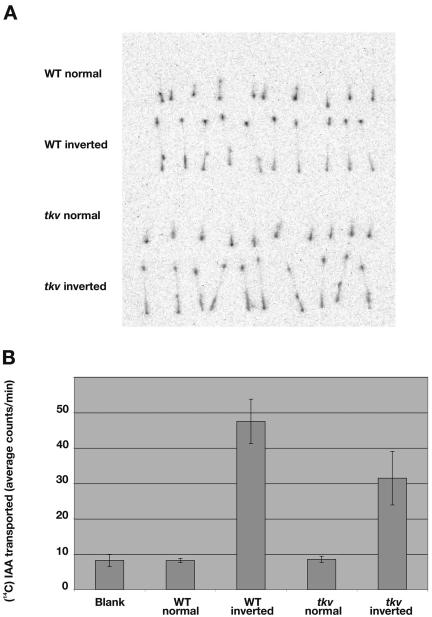
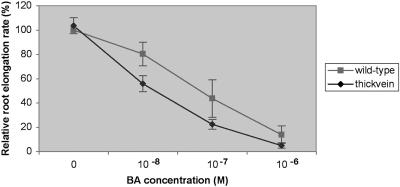
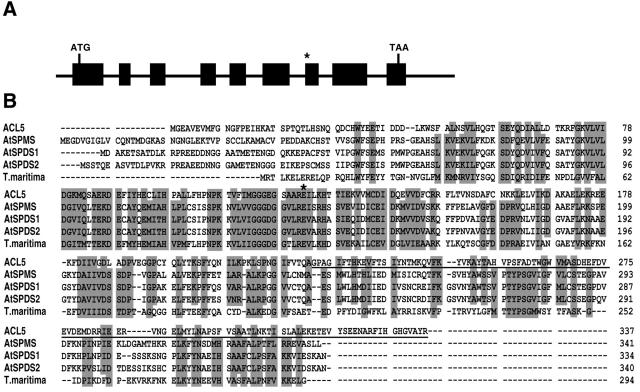
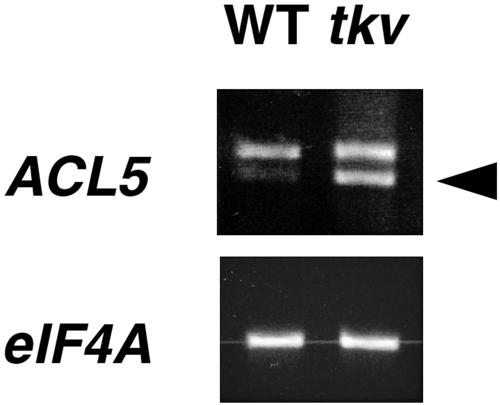
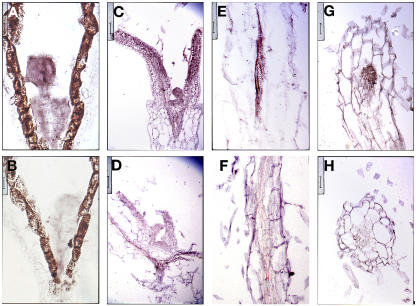
Polar auxin transport has been implicated in the induction of vascular tissue and in the definition of vein positions. Leaves treated with chemical inhibitors of polar auxin transport exhibited vascular phenotypes that include increased vein thickness and vascularization. We describe a recessive mutant, thickvein (tkv), which develops thicker veins in leaves and in inflorescence stems. The increased vein thickness is attributable to an increased number of vascular cells. Mutant plants have smaller leaves and shorter inflorescence stems, and this reduction in organ size and height is accompanied by an increase in organ vascularization, which appears to be attributable to an increase in the recruitment of cells into veins. Furthermore, although floral development is normal, auxin transport in the inflorescence stem is significantly reduced in the mutant, suggesting that the defect in auxin transport is responsible for the vascular phenotypes. In the primary root, the veins appear morphologically normal, but root growth in the tkv mutant is hypersensitive to exogenous cytokinin. The tkv mutation was found to reside in the ACL5 gene, which encodes a spermine synthase and whose expression is specific to provascular cells. We propose that ACL5/TKV is involved in vein definition (defining the boundaries between veins and nonvein regions) and in polar auxin transport, and that polyamines are involved in this process.
Figures
References
-
- Aloni R (2001) Foliar and axial aspects of vascular differentiation: hypotheses and evidence. J Plant Growth Regul 20: 22–34
-
- Antognoni F, Fornale S, Grimmer C, Komor E, Bagni N (1998) Long-distance translocation of polyamines in phloem and xylem of Ricinus communis L. plants. Planta 204: 520–527
-
- Bell CJ, Ecker JR (1993) Assignment of 30 microsatellite loci to the linkage map of Arabidopsis. Genomics 19: 137–144 - PubMed
-
- Benjamins R, Quint A, Weijers D, Hooykaas P, Offringa R (2001) The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport. Development 128: 4057–4067 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
