

Smad1 and Smad8 function similarly in mammalian central nervous system development
- PMID: 15899870
- PMCID: PMC1140628
- DOI: 10.1128/MCB.25.11.4683-4692.2005
Smad1 and Smad8 function similarly in mammalian central nervous system development
Abstract
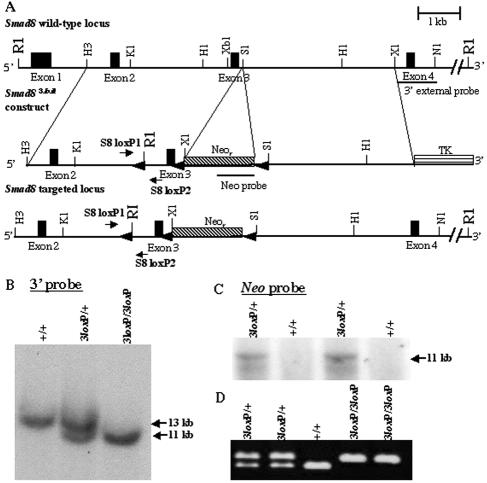
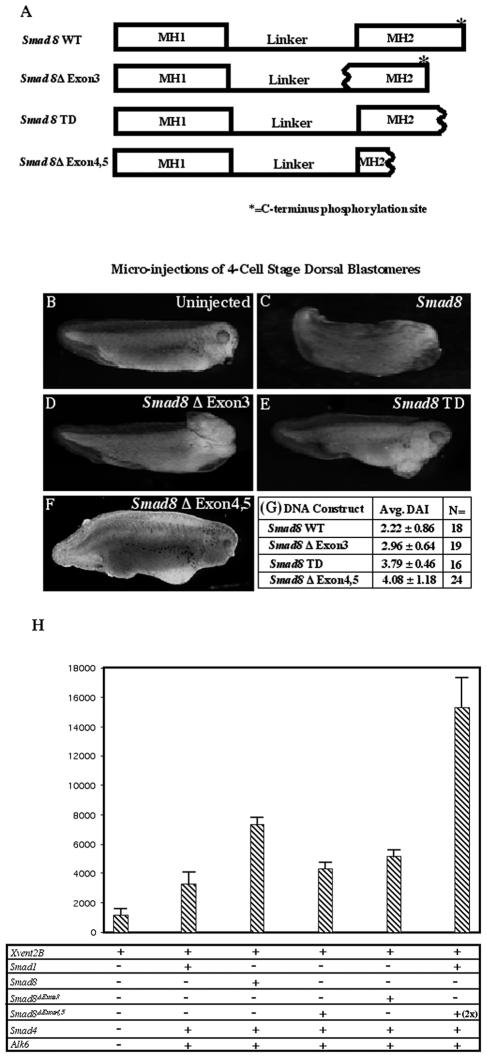
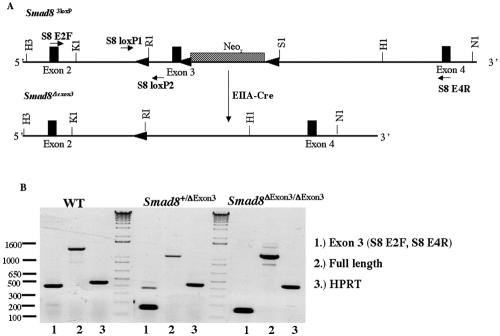
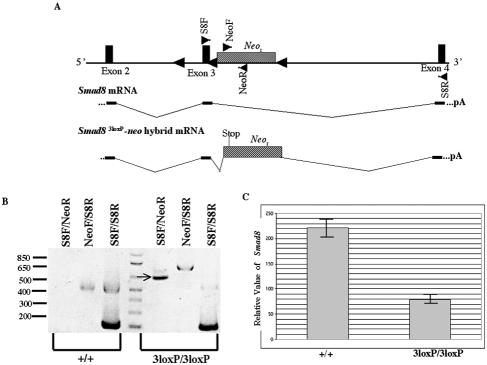
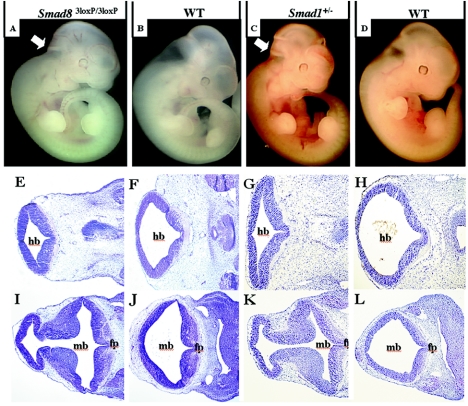
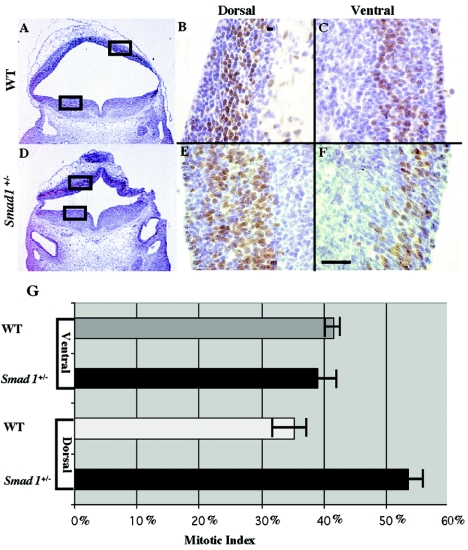
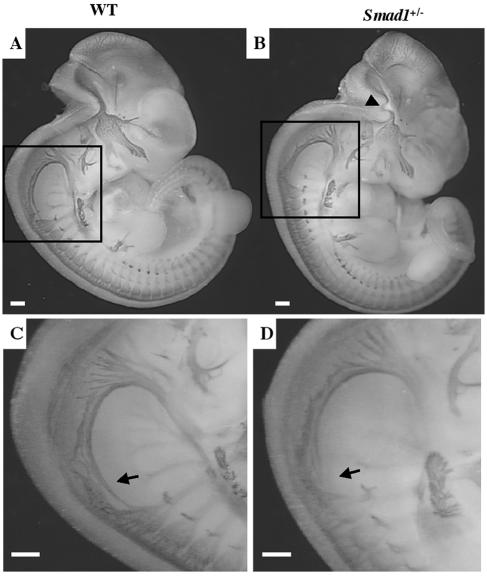
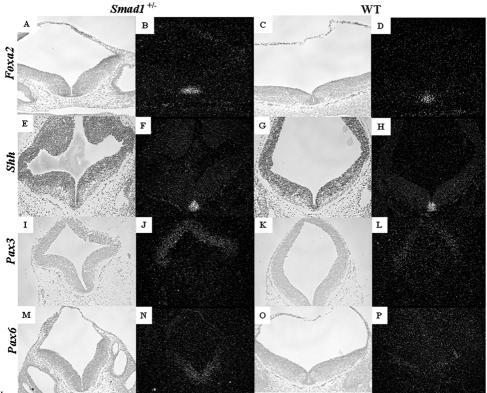
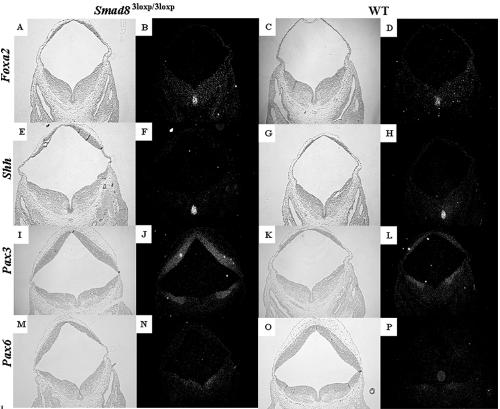
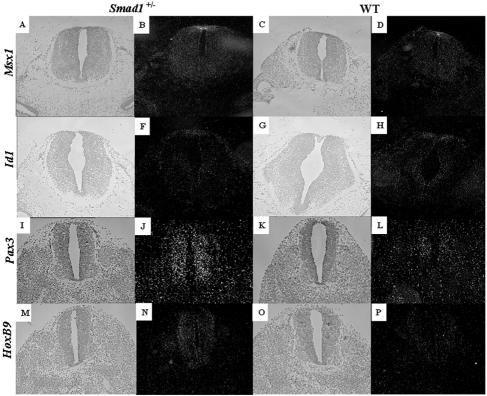
Smads 1, 5, and 8 are the intracellular mediators for the bone morphogenetic proteins (BMPs), which play crucial roles during mammalian development. Previous research has shown that Smad1 is important in the formation of the allantois, while Smad5 has been shown to be critical in the process of angiogenesis. To further analyze the BMP-responsive Smads, we disrupted the murine Smad8 gene utilizing the Cre/loxP system. A Smad8 hypomorphic allele (Smad8(Deltaexon3)) was constructed that contains an in-frame deletion of exon 3, removing one-third of the MH2 domain and a small portion of the linker region. Xenopus injection assays indicated that this Smad8 deletion allele is still functional but has reduced ventralizing capability compared to the wild type. Although Smad8(Deltaexon3/Deltaexon3) embryos are phenotypically normal, homozygotes of another hypomorphic allele of Smad8 (Smad8(3loxP)) containing a neomycin cassette within intron 3, phenocopy an embryonic brain defect observed in roughly 22% of Smad1(+/)(-) embryos analyzed at embryonic day 11.5. These observations suggest that BMP-responsive Smads have critical functions in the development of the mammalian central nervous system.
Figures
Similar articles
-
[The role of Smads and related transcription factors in the signal transduction of bone morphogenetic protein inducing bone formation].Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2003 Sep;17(5):359-62. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2003. PMID: 14551929 Review. Chinese.
-
Xenopus Smad8 acts downstream of BMP-4 to modulate its activity during vertebrate embryonic patterning.Development. 1998 Mar;125(5):857-67. doi: 10.1242/dev.125.5.857. Development. 1998. PMID: 9449668
-
BMP-Smad 1/5/8 signalling in the development of the nervous system.Prog Neurobiol. 2013 Oct;109:28-41. doi: 10.1016/j.pneurobio.2013.07.002. Epub 2013 Jul 24. Prog Neurobiol. 2013. PMID: 23891815 Review.
-
Bone morphogenetic protein-mediating receptor-associated Smads as well as common Smad are expressed in human articular chondrocytes but not up-regulated or down-regulated in osteoarthritic cartilage.J Bone Miner Res. 2002 Dec;17(12):2141-50. doi: 10.1359/jbmr.2002.17.12.2141. J Bone Miner Res. 2002. PMID: 12469908
-
Mouse embryos lacking Smad1 signals display defects in extra-embryonic tissues and germ cell formation.Development. 2001 Sep;128(18):3609-21. doi: 10.1242/dev.128.18.3609. Development. 2001. PMID: 11566864
Cited by
-
The multiple activities of BMPs during spinal cord development.Cell Mol Life Sci. 2013 Nov;70(22):4293-305. doi: 10.1007/s00018-013-1354-9. Epub 2013 May 15. Cell Mol Life Sci. 2013. PMID: 23673983 Free PMC article. Review.
-
Co-ordinating Notch, BMP, and TGF-β signaling during heart valve development.Cell Mol Life Sci. 2013 Aug;70(16):2899-917. doi: 10.1007/s00018-012-1197-9. Epub 2012 Nov 16. Cell Mol Life Sci. 2013. PMID: 23161060 Free PMC article. Review.
-
Bu-M-P-ing Iron: How BMP Signaling Regulates Muscle Growth and Regeneration.J Dev Biol. 2020 Feb 11;8(1):4. doi: 10.3390/jdb8010004. J Dev Biol. 2020. PMID: 32053985 Free PMC article. Review.
-
Inactivation of Smad5 in endothelial cells and smooth muscle cells demonstrates that Smad5 is required for cardiac homeostasis.Am J Pathol. 2007 May;170(5):1460-72. doi: 10.2353/ajpath.2007.060839. Am J Pathol. 2007. PMID: 17456754 Free PMC article.
-
The MH1 domain of Smad3 interacts with Pax6 and represses autoregulation of the Pax6 P1 promoter.Nucleic Acids Res. 2007;35(3):890-901. doi: 10.1093/nar/gkl1105. Epub 2007 Jan 23. Nucleic Acids Res. 2007. PMID: 17251190 Free PMC article.
References
-
- Anderson, R. M., A. R. Lawrence, R. W. Stottmann, D. Bachiller, and J. Klingensmith. 2002. Chordin and noggin promote organizing centers of forebrain development in the mouse. Development 129:4975-4987. - PubMed
-
- Ang, S. L., and J. Rossant. 1994. HNF-3 beta is essential for node and notochord formation in mouse development. Cell 78:561-574. - PubMed
-
- Attisano, L., and J. L. Wrana. 2002. Signal transduction by the TGF-beta superfamily. Science 296:1646-1647. - PubMed
-
- Bachiller, D., J. Klingensmith, C. Kemp, J. A. Belo, R. M. Anderson, S. R. May, J. A. McMahon, A. P. McMahon, R. M. Harland, J. Rossant, and E. M. De Robertis. 2000. The organizer factors Chordin and Noggin are required for mouse forebrain development. Nature 403:658-661. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases