

Fructose utilization in Lactococcus lactis as a model for low-GC gram-positive bacteria: its regulator, signal, and DNA-binding site
- PMID: 15901699
- PMCID: PMC1112048
- DOI: 10.1128/JB.187.11.3752-3761.2005
Fructose utilization in Lactococcus lactis as a model for low-GC gram-positive bacteria: its regulator, signal, and DNA-binding site
Abstract
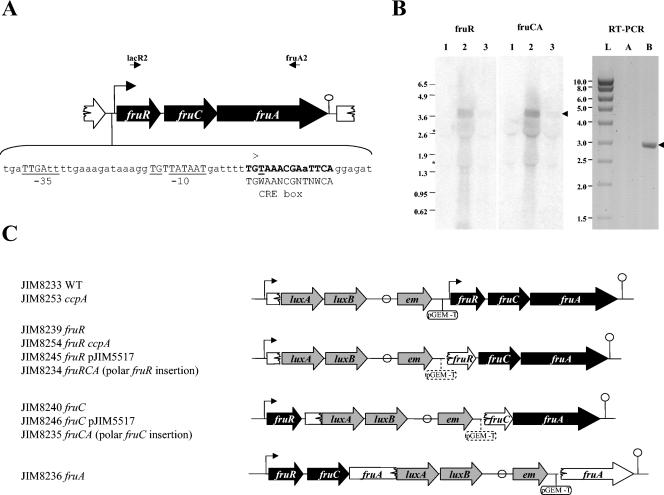
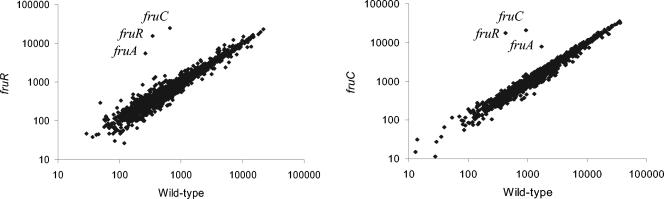
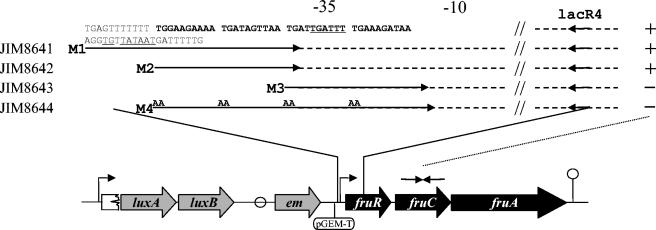
In addition to its role as carbon and energy source, fructose metabolism was reported to affect other cellular processes, such as biofilm formation by streptococci and bacterial pathogenicity in plants. Fructose genes encoding a 1-phosphofructokinase and a phosphotransferase system (PTS) fructose-specific enzyme IIABC component reside commonly in a gene cluster with a DeoR family regulator in various gram-positive bacteria. We present a comprehensive study of fructose metabolism in Lactococcus lactis, including a systematic study of fru mutants, global messenger analysis, and a molecular characterization of its regulation. The fru operon is regulated at the transcriptional level by both FruR and CcpA and at the metabolic level by inducer exclusion. The FruR effector is fructose-1-phosphate (F1P), as shown by combined analysis of transcription and measurements of the intracellular F1P pools in mutants either unable to produce this metabolite or accumulating it. The regulation of the fru operon by FruR requires four adjacent 10-bp direct repeats. The well-conserved organization of the fru promoter region in various low-GC gram-positive bacteria, including CRE boxes as well as the newly defined FruR motif, suggests that the regulation scheme defined in L. lactis could be applied to these bacteria. Transcriptome profiling of fruR and fruC mutants revealed that the effect of F1P and FruR regulation is limited to the fru operon in L. lactis. This result is enforced by the fact that no other targets for FruR were found in the available low-GC gram-positive bacteria genomes, suggesting that additional phenotypical effects due to fructose metabolism do not rely directly on FruR control, but rather on metabolism.
Figures

References
-
- Bailey, T. L., and C. Elkan. 1995. The value of prior knowledge in discovering motifs with MEME. Proc. Int. Conf. Intell. Syst. Mol. Biol. 3:21-29. - PubMed
-
- Brennan, R. G., and B. W. Matthews. 1989. Structural basis of DNA-protein recognition. Trends Biochem. Sci. 14:286-290. - PubMed
-
- Bruckner, R., and F. Titgemeyer. 2002. Carbon catabolite repression in bacteria: choice of the carbon source and autoregulatory limitation of sugar utilization. FEMS Microbiol. Lett. 209:141-148. - PubMed
-
- Chiou, C. Y., H. H. Wang, and G. C. Shaw. 2002. Identification and characterization of the non-PTS fru locus of Bacillus megaterium ATCC 14581. Mol. Genet. Genomics 268:240-248. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
