

Synaptic transmission at the cochlear nucleus endbulb synapse during age-related hearing loss in mice
- PMID: 15901757
- PMCID: PMC1941703
- DOI: 10.1152/jn.00374.2005
Synaptic transmission at the cochlear nucleus endbulb synapse during age-related hearing loss in mice
Abstract
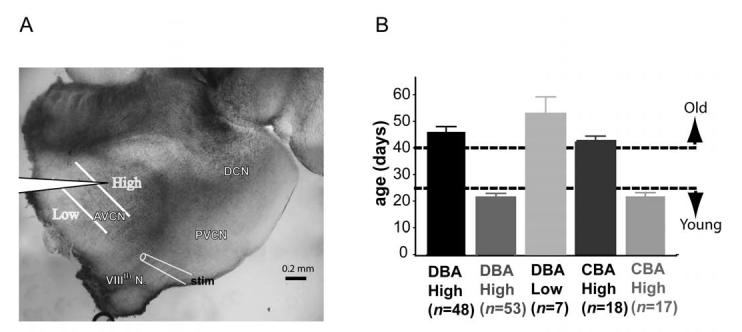
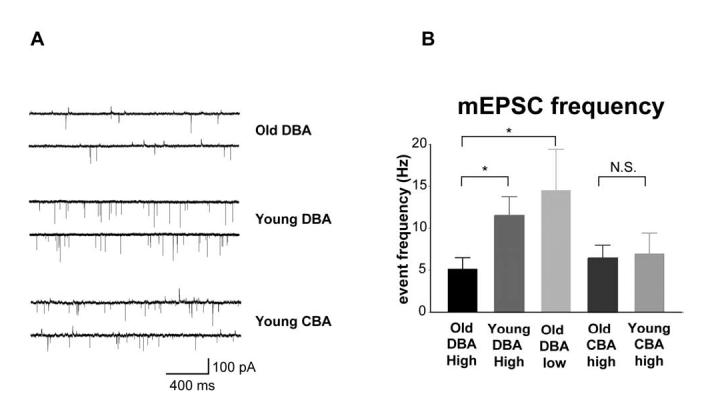
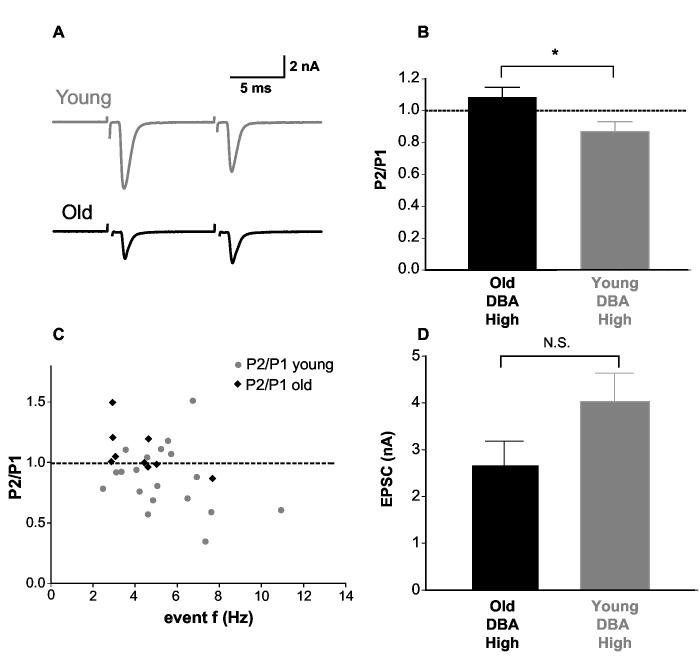
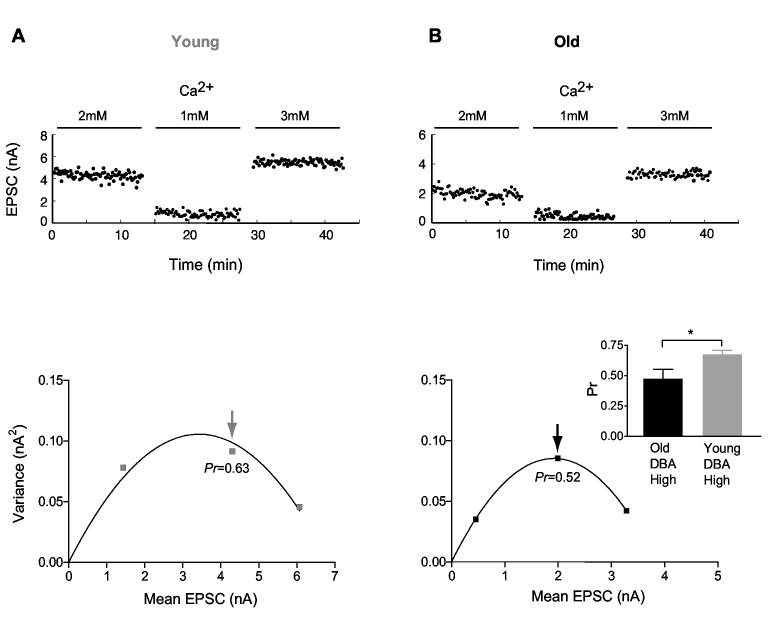
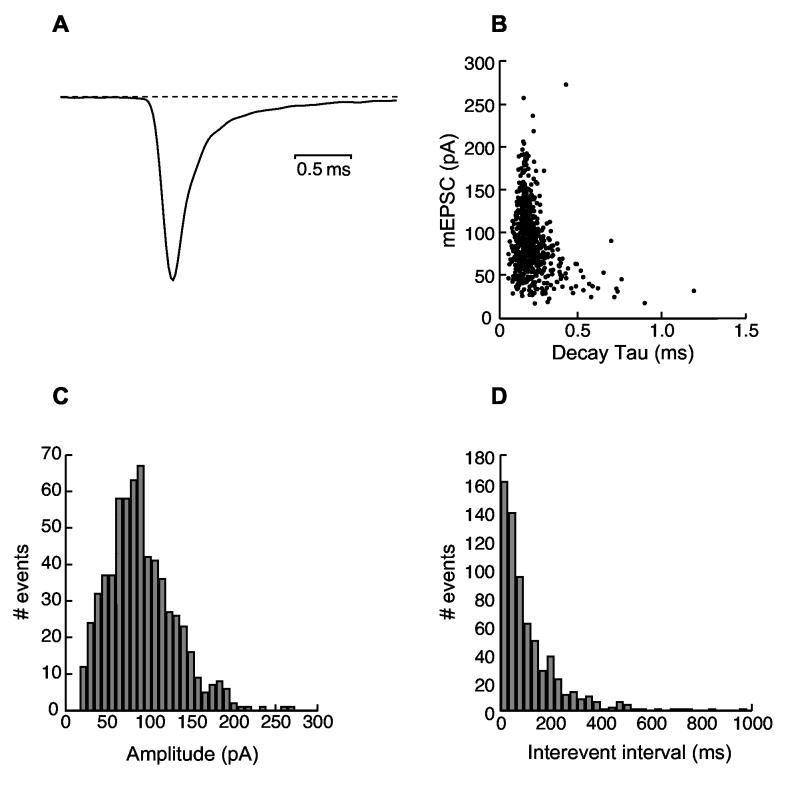
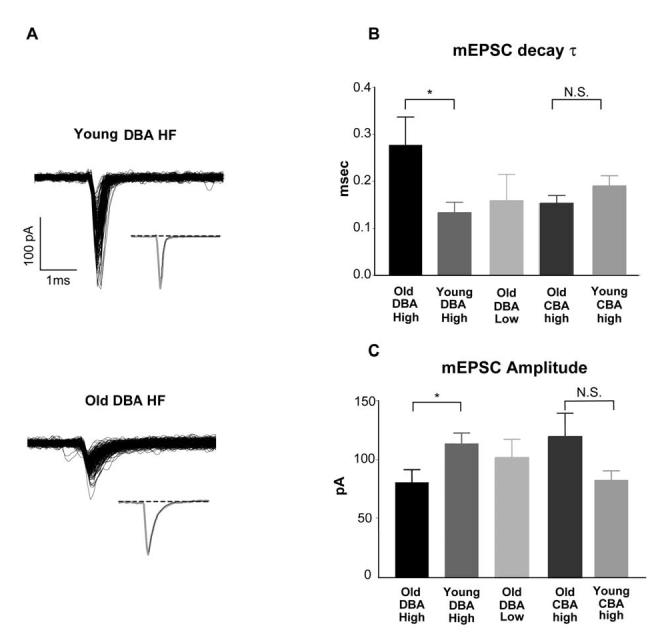
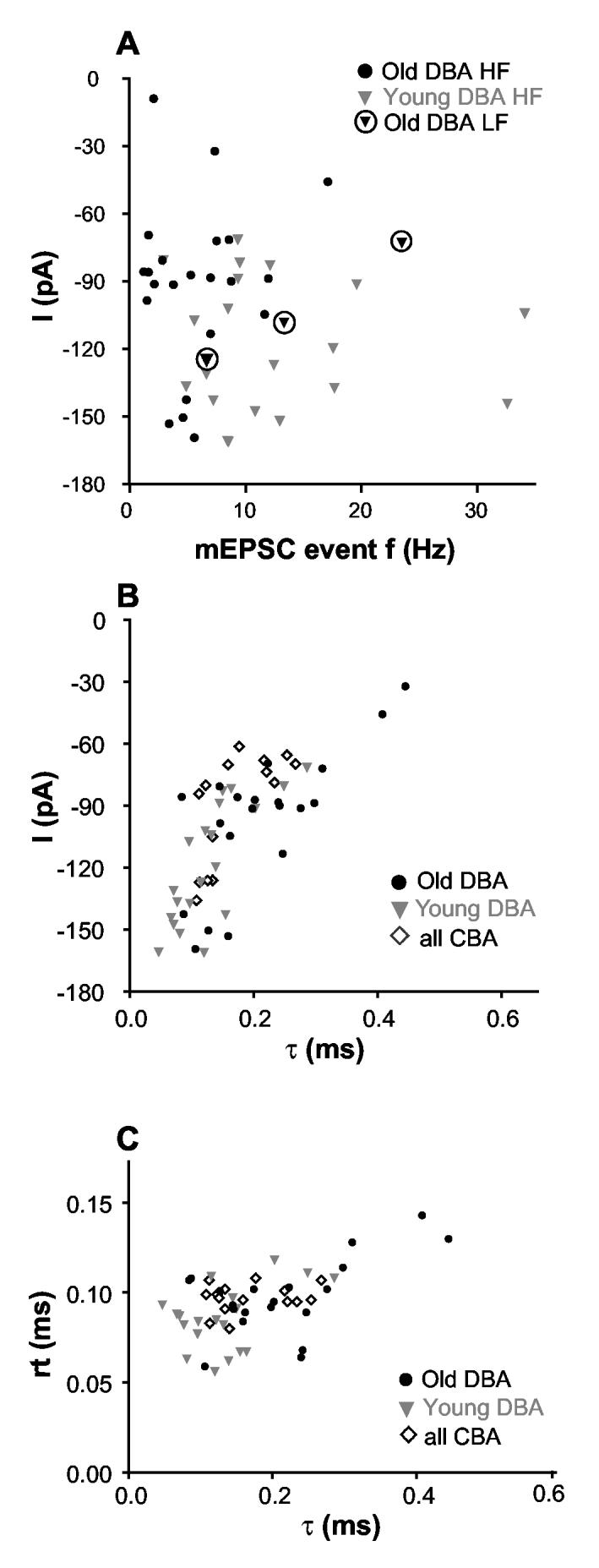
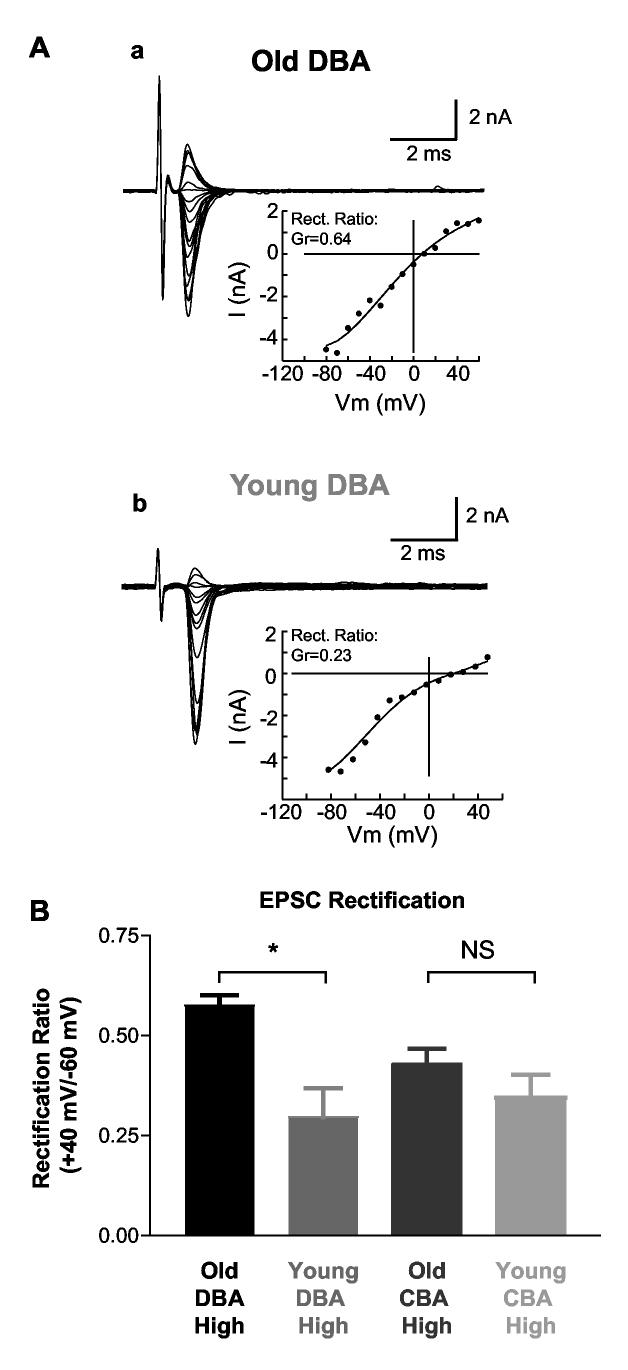
Age-related hearing loss (AHL) typically starts from high-frequency regions of the cochlea and over time invades lower-frequency regions. During this progressive hearing loss, sound-evoked activity in spiral ganglion cells is reduced. DBA mice have an early onset of AHL. In this study, we examined synaptic transmission at the endbulb of Held synapse between auditory nerve fibers and bushy cells in the anterior ventral cochlear nucleus (AVCN). Synaptic transmission in hearing-impaired high-frequency areas of the AVCN was altered in old DBA mice. The spontaneous miniature excitatory postsynaptic current (mEPSC) frequency was substantially reduced (about 60%), and mEPSCs were significantly slower (about 115%) and smaller (about 70%) in high-frequency regions of old (average age 45 days) DBA mice compared with tonotopically matched regions of young (average age 22 days) DBA mice. Moreover, synaptic release probability was about 30% higher in high-frequency regions of young DBA than that in old DBA mice. Auditory nerve-evoked EPSCs showed less rectification in old DBA mice, suggesting recruitment of GluR2 subunits into the AMPA receptor complex. No similar age-related changes in synaptic release or EPSCs were found in age-matched, normal hearing young and old CBA mice. Taken together, our results suggest that auditory nerve activity plays a critical role in maintaining normal synaptic function at the endbulb of Held synapse after the onset of hearing. Auditory nerve activity regulates both presynaptic (release probability) and postsynaptic (receptor composition and kinetics) function at the endbulb synapse after the onset of hearing.
Figures
References
-
- Berglund AM, Brown MC. Central trajectories of type II spiral ganglion cells from various cochlear regions in mice. Hear Res. 1994;75:121–130. - PubMed
-
- Boulter J, Hollmann M, O’Shea-Greenfield A, Hartley M, Deneris E, Maron C, Heinemann S. Molecular cloning and functional expression of glutamate receptor subunit genes. Science. 1990;249:1033–1037. - PubMed
-
- Bowie D, Mayer ML. Inward rectification of both AMPA and kainate subtype glutamate receptors generated by polyamine-mediated ion channel block. Neuron. 1995;15:453–462. - PubMed
-
- Brawer JR, Morest DK, Kane EC. The neuronal architecture of the cochlear nucleus of the cat. J Comp Neurol. 1974;155:251–300. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical