

Atg17 regulates the magnitude of the autophagic response
- PMID: 15901835
- PMCID: PMC1165424
- DOI: 10.1091/mbc.e04-10-0894
Atg17 regulates the magnitude of the autophagic response
Abstract
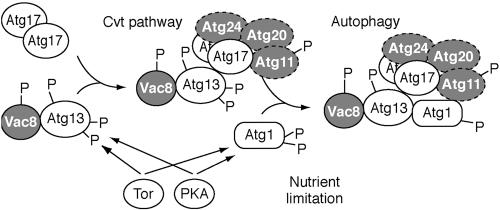
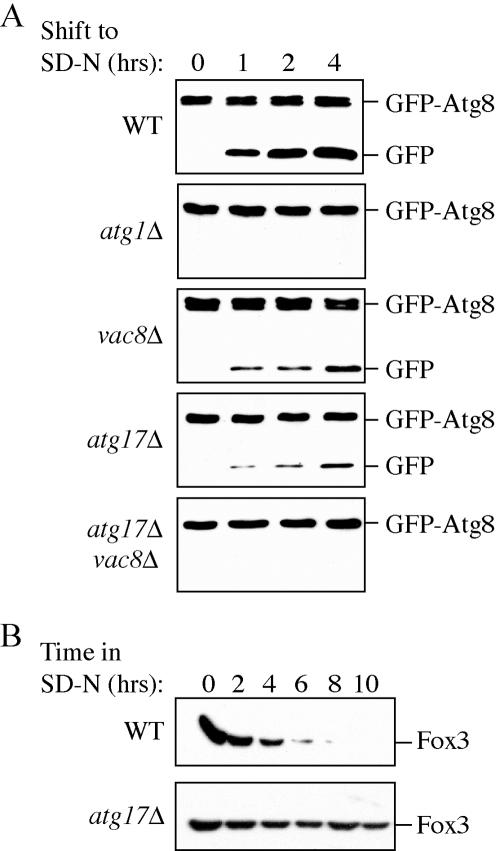
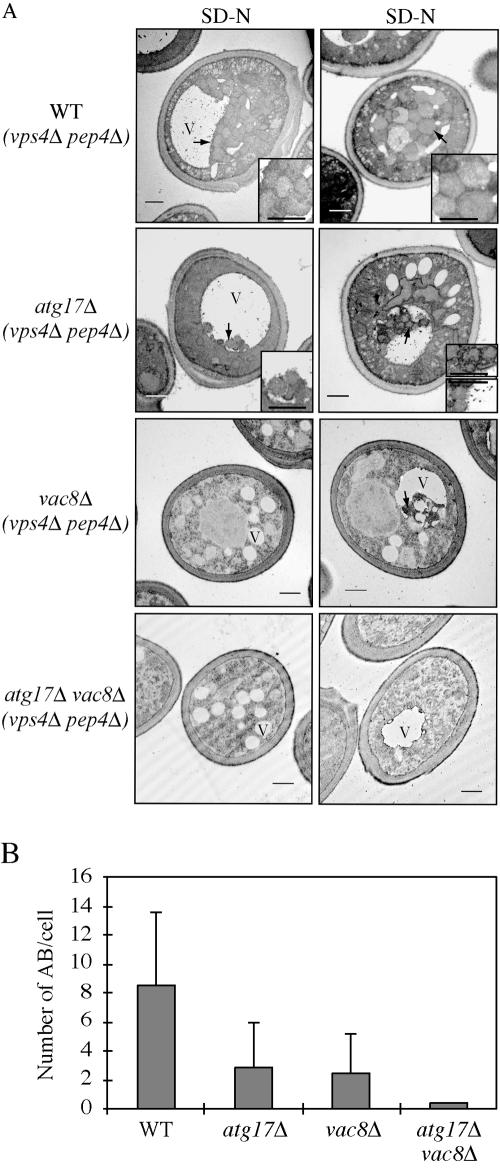
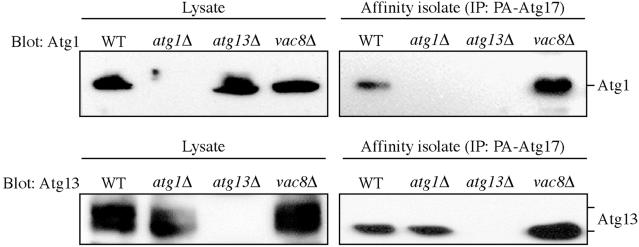
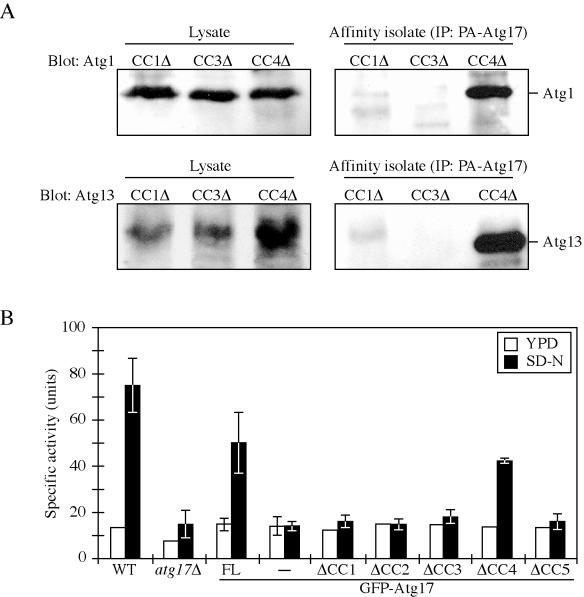
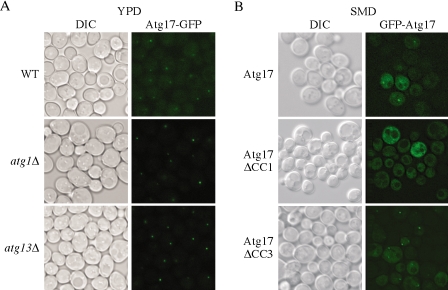
Autophagy is a catabolic process used by eukaryotic cells for the degradation and recycling of cytosolic proteins and excess or defective organelles. In yeast, autophagy is primarily a response to nutrient limitation, whereas in higher eukaryotes it also plays a role in developmental processes. Due to its essentially unlimited degradative capacity, it is critical that regulatory mechanisms are in place to modulate the timing and magnitude of the autophagic response. One set of proteins that seems to function in this regard includes a complex that contains the Atg1 kinase. Aside from Atg1, the proteins in this complex participate primarily in either nonspecific autophagy or specific types of autophagy, including the cytoplasm to vacuole targeting pathway, which operates under vegetative growth conditions, and peroxisome degradation. Accordingly, these proteins are prime candidates for factors that regulate the conversion between these pathways, including the change in size of the sequestering vesicle, the most obvious morphological difference. The atg17delta mutant forms a reduced number of small autophagosomes. As a result, it is defective in peroxisome degradation and is partially defective for autophagy. Atg17 interacts with both Atg1 and Atg13, via two coiled-coil domains, and these interactions facilitate its inclusion in the Atg1 complex.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
