

Non-equilibration of hydrostatic pressure in blebbing cells
- PMID: 15902261
- PMCID: PMC1564437
- DOI: 10.1038/nature03550
Non-equilibration of hydrostatic pressure in blebbing cells
Abstract
Current models for protrusive motility in animal cells focus on cytoskeleton-based mechanisms, where localized protrusion is driven by local regulation of actin biochemistry. In plants and fungi, protrusion is driven primarily by hydrostatic pressure. For hydrostatic pressure to drive localized protrusion in animal cells, it would have to be locally regulated, but current models treating cytoplasm as an incompressible viscoelastic continuum or viscous liquid require that hydrostatic pressure equilibrates essentially instantaneously over the whole cell. Here, we use cell blebs as reporters of local pressure in the cytoplasm. When we locally perfuse blebbing cells with cortex-relaxing drugs to dissipate pressure on one side, blebbing continues on the untreated side, implying non-equilibration of pressure on scales of approximately 10 microm and 10 s. We can account for localization of pressure by considering the cytoplasm as a contractile, elastic network infiltrated by cytosol. Motion of the fluid relative to the network generates spatially heterogeneous transients in the pressure field, and can be described in the framework of poroelasticity.
Figures
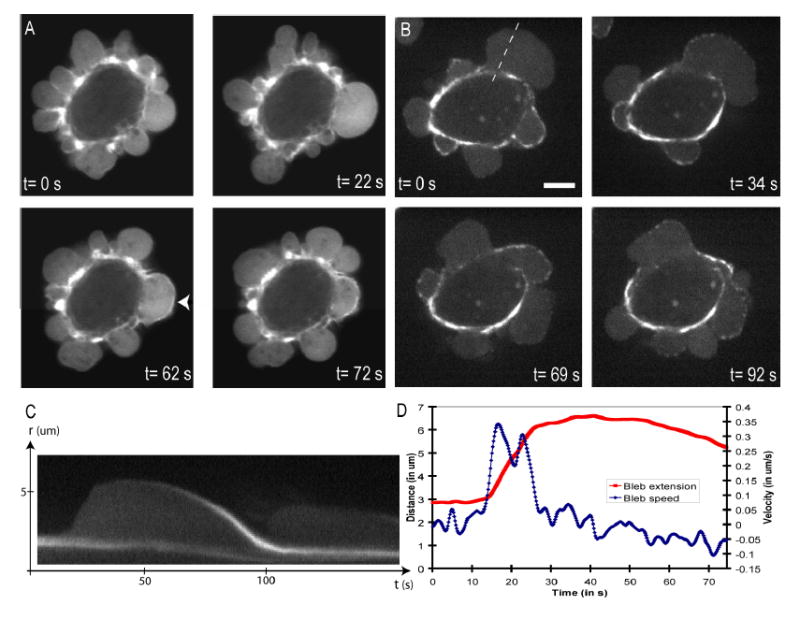
GFP-actin transfected cell. During bleb expansion, the bleb rim does not appear enriched in GFP-actin compared to the bleb interior (t=0s and t=22s). Once expansion has halted, an actin-rich rim forms at the bleb surface (t=62s, arrow head) and retraction begins. As retraction proceeds, the actin rim becomes wavy, suggesting that the cortex contracts as the bleb shrinks (t=72s).
GFP-MRLC transfected cell. Just after expansion has stopped, the bleb surface does not appear to be enriched in GFP-MRLC (t=0s). MRLC then accumulates in discrete foci (t=34s) and retraction starts (t=69s). As retraction ends, MRLC forms a continuum along the bleb rim (t=92s).
Kymograph of the expansion and retraction of the bleb in 1B (dashed line). As the bleb expands, there is no accumulation of MRLC at the bleb apex. As retraction proceeds, MRLC accumulates at the bleb.
Graph of the bleb extension and bleb velocity as a function of time for the bleb shown in 1C. The maximal bleb extension is ~3.5 μm (maximal speed of expansion ~0.35 μm.s−1, speed of retraction ~0.1 μm.s−1).
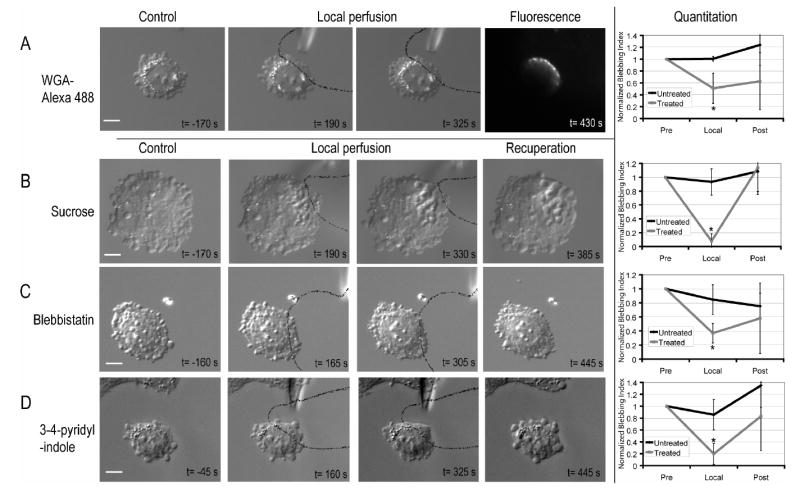
Local perfusion of Wheat Germ Agglutinin (WGA) –Alexa 488. WGA-Alexa 488 was only incorporated in the region of the cell that was exposed to flow from the micropipette.
Local perfusion of a 300mM sucrose solution. Vacuoles form in the vicinity of the perfused region.
Local inhibition of myosin II ATP-ase by local perfusion of blebbistatin
Local inhibition of ROCKI by 3-(4-pyridyl)indole. Solid black lines delineate the flow out of the micropipette. On each image, the inset text gives timing relative to local application of treatment. The normalised blebbing indices show the evolution over time of blebbing in the region exposed to inhibitor and in the free region. Error bars show the standard deviation. Asterisks denote significant changes in the blebbing index when compared to the initial blebbing index. Scale bars=10μm.

Local perfusion with Latrunculin B has a dual effect. When Latrunculin B is applied locally, bleb size increases globally (t=75s), and bleb dynamics cease in the treated area (t=75s, t=315s). Elsewhere, expansion and retraction continue.
Global perfusion of Staurosporine is necessary to inhibit blebbing. Prior to perfusion (t=−170s) and after 395 s of local perfusion with Staurosporine, blebbing is unperturbed throughout the cell. When Staurosporine is applied to the whole cell (white letters, t=450s), blebbing ceases.
Global perfusion of HA-1077 is necessitated to inhibit blebbing. After 340s of local HA-1077 application, blebbing is unperturbed. When the whole cell is exposed to inhibitor (t=230s, white letters), blebbing ceases. When perfusion is halted, blebbing is restored (t=535s, white letters).
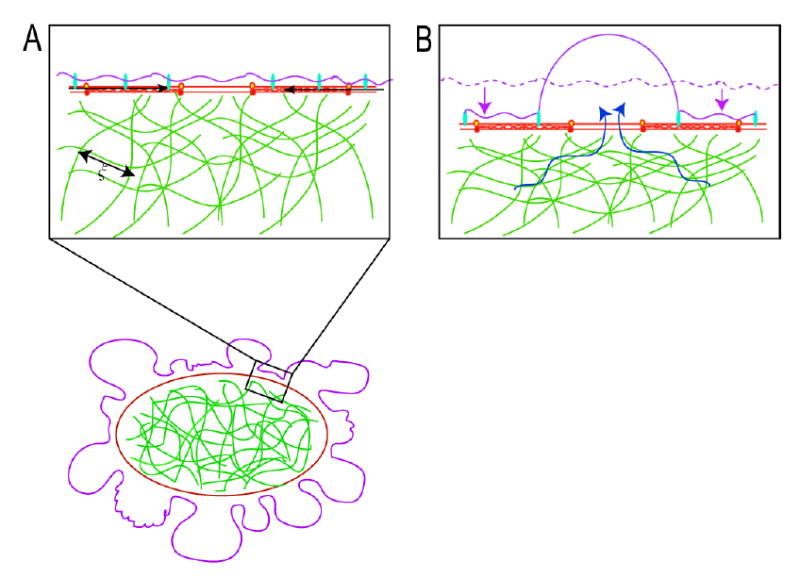
In blebbing cells, a local contraction of the myosin II (black arrows) associated with the actin cortex leads to a shortening of the cortical periphery and therefore to a compression of the cytoskeletal network that fills the cell. The cytoskeletal network is porous and has an average pore size ξ. The compression of the cytoskeletal meshwork creates a hydrostatic pressure in the vicinity of the region of contraction and can lead to bleb nucleation and expansion.
Contraction of the actin cortex leads to a compression of the cytoskeletal network (the dashed mauve line indicates the original position of the cell surface) drives flow of cytosol in the opposite direction (blue arrows). If there is a local defect in membrane-cytoskeleton attachment, a bleb is extruded. Bleb expansion is opposed by two forces: extracellular osmotic pressure and membrane tension.
References
-
- Mahadevan L, Matsudaira P. Motility powered by supramolecular springs and ratchets. Science. 2000;288:95–100. - PubMed
-
- Rafelski SM, Theriot JA. Crawling toward a unified model of cell mobility: spatial and temporal regulation of actin dynamics. Annu Rev Biochem. 2004;73:209–39. - PubMed
-
- Mogilner A, Oster G. Polymer motors: pushing out the front and pulling up the back. Curr Biol. 2003;13:R721–33. - PubMed
-
- Messerli MA, Robinson KR. Ionic and osmotic disruptions of the lily pollen tube oscillator: testing proposed models. Planta. 2003;217:147–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
