

Morphogenesis of the right ventricle requires myocardial expression of Gata4
- PMID: 15902305
- PMCID: PMC1090473
- DOI: 10.1172/JCI23769
Morphogenesis of the right ventricle requires myocardial expression of Gata4
Abstract
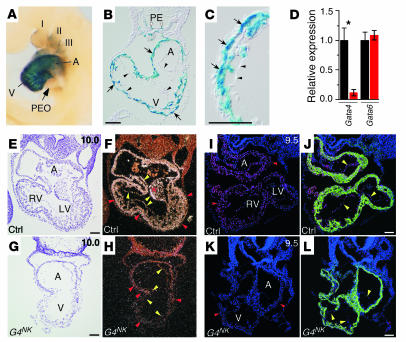
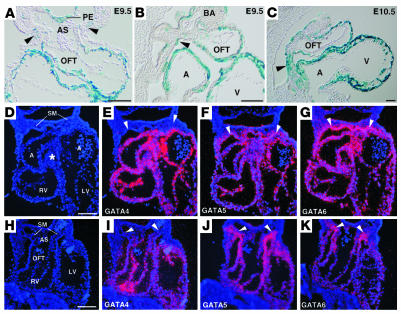
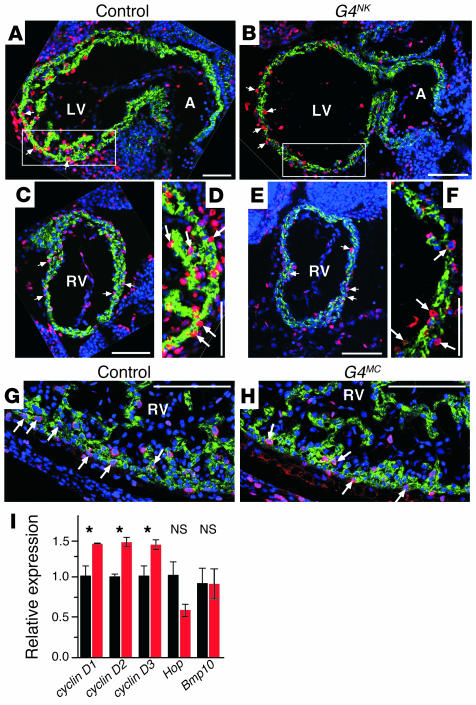
Mutations in developmental regulatory genes have been found to be responsible for some cases of congenital heart defects. One such regulatory gene is Gata4, a zinc finger transcription factor. In order to circumvent the early embryonic lethality of Gata4-null embryos and to investigate the role of myocardial Gata4 expression in cardiac development, we used Cre/loxP technology to conditionally delete Gata4 in the myocardium of mice at an early and a late time point in cardiac morphogenesis. Early deletion of Gata4 by Nkx2-5Cre resulted in hearts with striking myocardial thinning, absence of mesenchymal cells within the endocardial cushions, and selective hypoplasia of the RV. RV hypoplasia was associated with downregulation of Hand2, a transcription factor previously shown to regulate formation of the RV. Cardiomyocyte proliferation was reduced, with a greater degree of reduction in the RV than in the LV. Late deletion of Gata4 by Cre recombinase driven by the alpha myosin heavy chain promoter did not selectively affect RV development or generation of endocardial cushion mesenchyme but did result in marked myocardial thinning with decreased cardiomyocyte proliferation, as well as double-outlet RV. Our results demonstrate a general role of myocardial Gata4 in regulating cardiomyocyte proliferation and a specific, stage-dependent role in regulating the morphogenesis of the RV and the atrioventricular canal.
Figures




References
-
- Hoffman JI. Incidence of congenital heart disease: I. Postnatal incidence. Pediatr. Cardiol. 1995;16:103–113. - PubMed
-
- Waldo KL, et al. Conotruncal myocardium arises from a secondary heart field. Development. 2001;128:3179–3188. - PubMed
-
- Mjaatvedt CH, et al. The outflow tract of the heart is recruited from a novel heart-forming field. Dev. Biol. 2001;238:97–109. - PubMed
-
- Kelly RG, Brown NA, Buckingham ME. The arterial pole of the mouse heart forms from Fgf10-expressing cells in pharyngeal mesoderm. Dev. Cell. 2001;1:435–440. - PubMed
-
- Runyan RB, Markwald RR. Invasion of mesenchyme into three-dimensional collagen gels: a regional and temporal analysis of interaction in embryonic heart tissue. Dev. Biol. 1983;95:108–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
