

Entamoeba histolytica and Entamoeba dispar utilize externalized phosphatidylserine for recognition and phagocytosis of erythrocytes
- PMID: 15908370
- PMCID: PMC1111853
- DOI: 10.1128/IAI.73.6.3422-3430.2005
Entamoeba histolytica and Entamoeba dispar utilize externalized phosphatidylserine for recognition and phagocytosis of erythrocytes
Abstract
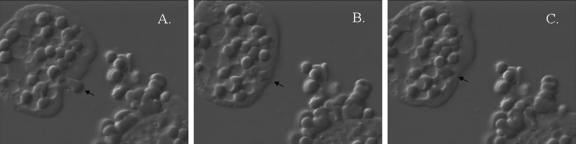
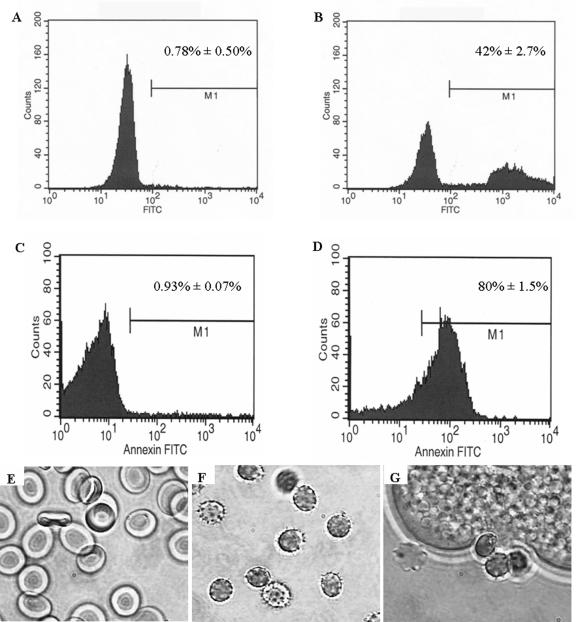
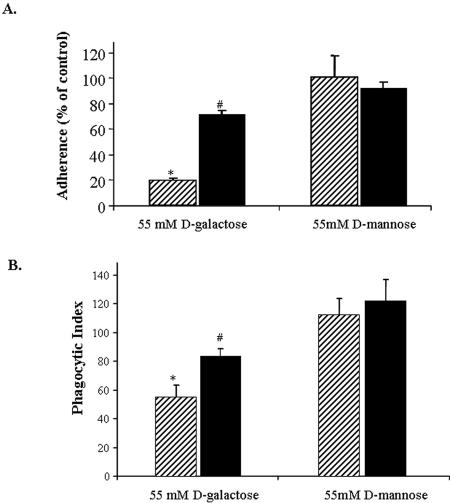
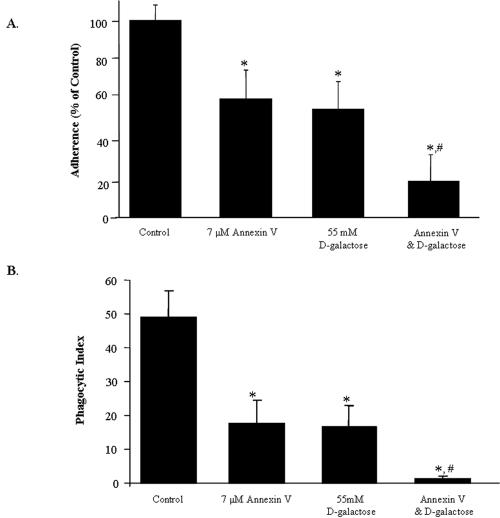
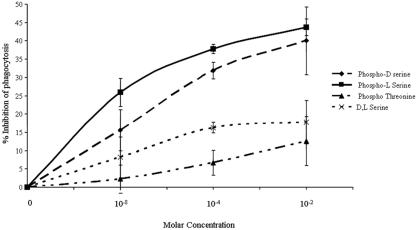
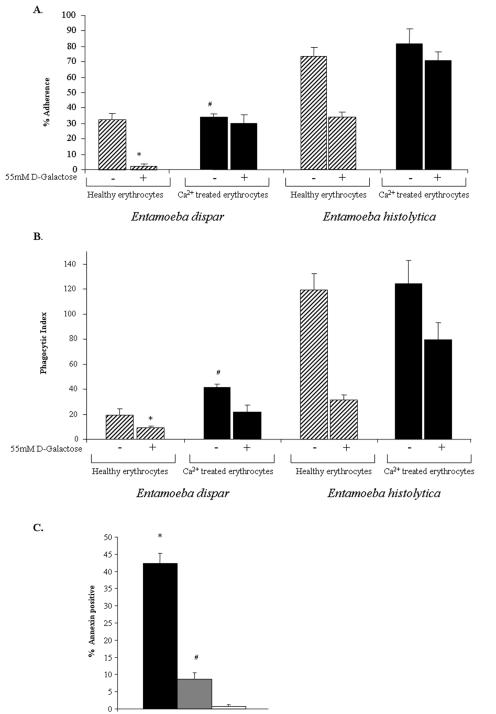
Amebic erythrophagocytosis is characteristic of invasive amebiasis, and mutants deficient in erythrocyte ingestion are avirulent. We sought to understand the molecular mechanisms underlying erythrocyte phagocytosis by Entamoeba histolytica. Following adherence to amebae, erythrocytes became round and crenulated, and phosphatidylserine (PS) was exposed on their outer membrane leaflets. These changes were similar to the effects of calcium treatment on erythrocytes, which we utilized to separate ameba-induced exposure of erythrocyte PS from the process of phagocytosis. The adherence and phagocytosis of calcium-treated erythrocytes were less inhibited by galactose than were those of healthy erythrocytes, suggesting the existence of an amebic coreceptor specific for PS. To test whether PS was recognized by amebae, calcium-treated cells were incubated with annexin V prior to adherence to or ingestion by E. histolytica. Annexin V blocked both adherence (50% +/- 12% inhibition; P < 0.05) and phagocytosis (65% +/- 10%; P < 0.05), providing evidence that at least one galactose-independent coreceptor was involved in the adherence and ingestion of red blood cells. The coreceptor was inhibited by phospho-l-serine and to a lesser extent by phospho-d-serine but not by phospho-l-threonine, which is consistent with the coreceptor functioning in the adherence and ingestion of erythrocytes via recognition of PS. We expanded our investigations to the highly related but noninvasive parasite Entamoeba dispar and demonstrated that it was deficient in red-blood-cell adherence, induction of PS exposure, and phagocytosis. These findings establish phosphatidylserine involvement in erythrophagocytosis by amebae and suggest the existence of a PS receptor on the surfaces of both E. histolytica and E. dispar.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
