

A theoretically estimated optimal cooling rate for the cryopreservation of sperm cells from a live-bearing fish, the green swordtail Xiphophorus helleri
- PMID: 15910922
- PMCID: PMC5592831
- DOI: 10.1016/j.theriogenology.2004.09.051
A theoretically estimated optimal cooling rate for the cryopreservation of sperm cells from a live-bearing fish, the green swordtail Xiphophorus helleri
Abstract
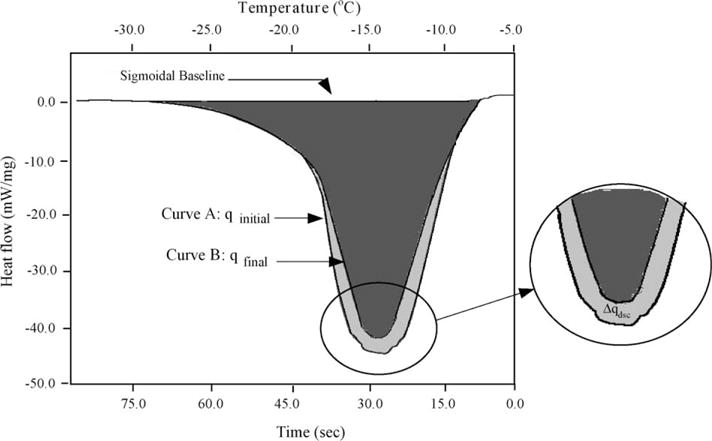
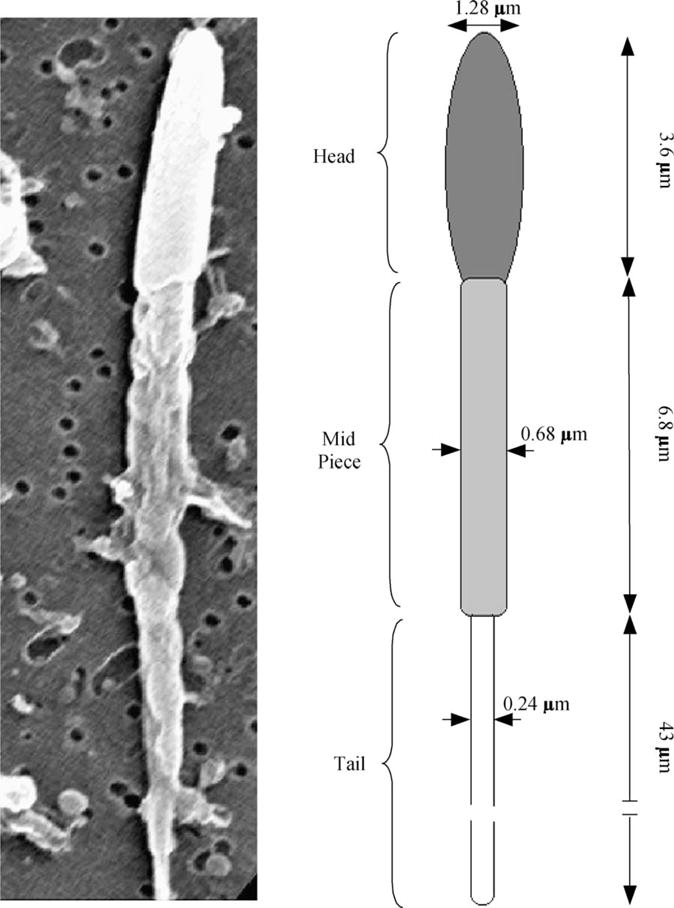
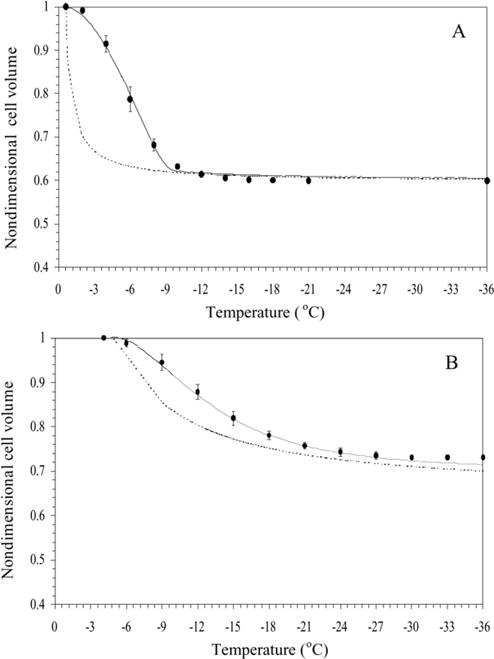
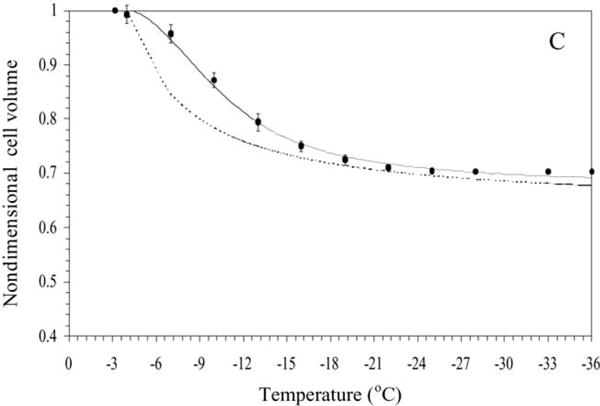
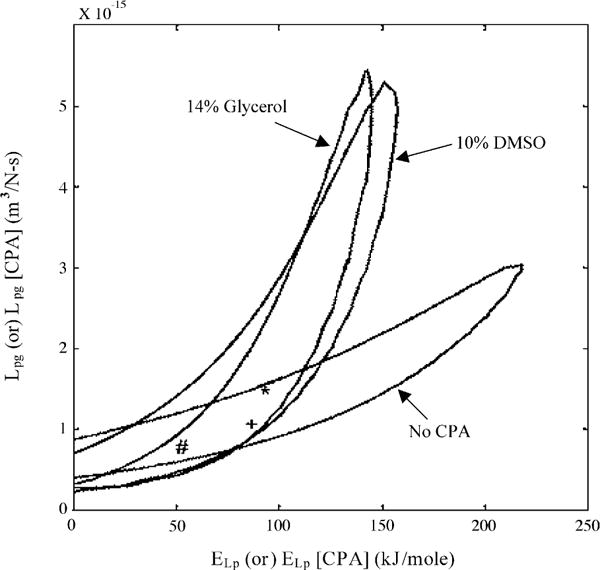
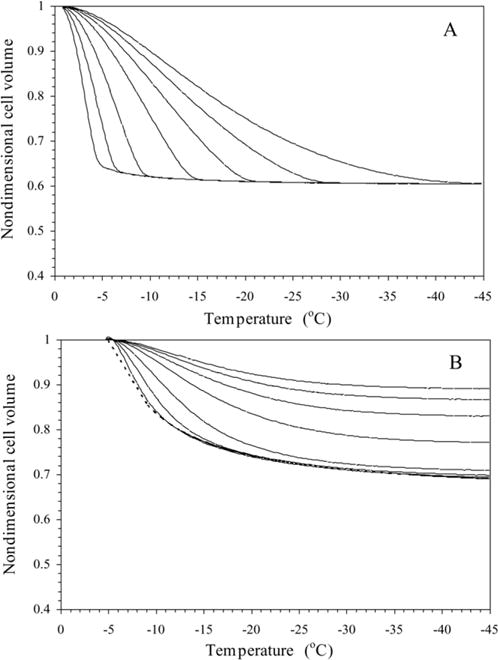
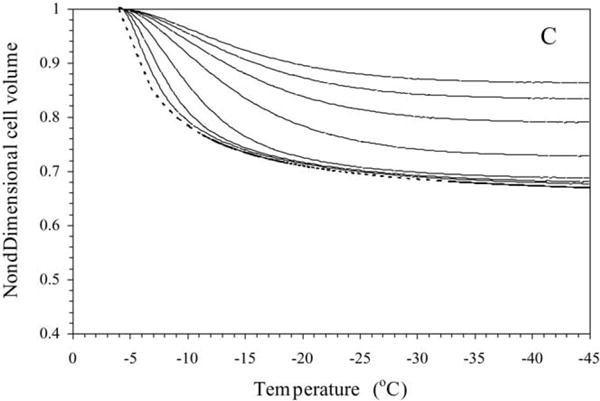
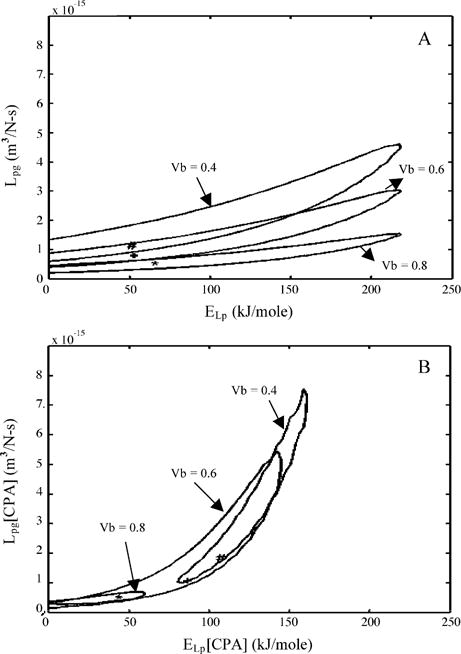
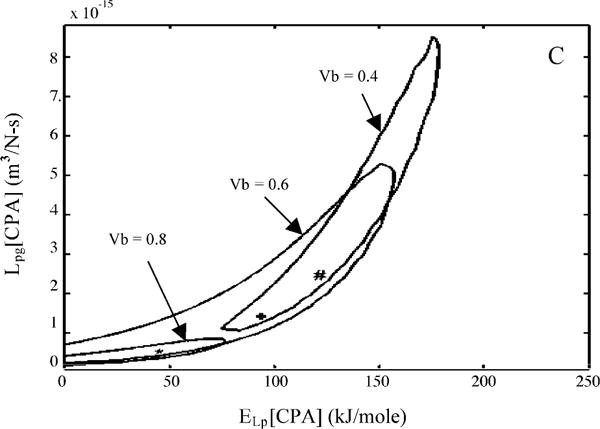
Sperm cryopreservation of live-bearing fishes, such as those of the genus Xiphophorus is only beginning to be studied, although these fishes are valuable models for biomedical research and are commercially raised as ornamental fish for use in aquariums. To explore optimization of techniques for sperm cryopreservation of these fishes, this study measured the volumetric shrinkage response during freezing of sperm cells of Xiphophorus helleri by use of a shape-independent differential scanning calorimeter (DSC) technique. Volumetric shrinkage during freezing of X. helleri sperm cell suspensions was obtained in the presence of extracellular ice at a cooling rate of 20 degrees C/min in three different media: (1) Hanks' balanced salt solution (HBSS) without cryoprotective agents (CPAs); (2) HBSS with 14% (v/v) glycerol; and (3) HBSS with 10% (v/v) dimethyl sulfoxide (DMSO). The sperm cell was modeled as a cylinder of 33.3 microm in length and 0.59 microm in diameter with an osmotically inactive cell volume (V(b)) of 0.6V(o), where V(o) is the isotonic or initial cell volume. By fitting a model of water transport to the experimentally determined volumetric shrinkage data, the best-fit membrane permeability parameters (reference membrane permeability to water, L(pg) or L(pg)[cpa] and the activation energy, E(Lp) or E(Lp)[cpa]) of the Xiphophorus helleri sperm cell membrane were determined. The best-fit membrane permeability parameters at 20 degrees C/min in the absence of CPAs were: L(pg)=0.776 x 10(-15)m3/Ns (0.0046 microm/min atm), and E(Lp)=50.1 kJ/mol (11.97 kcal/mol) (R2=0.997). The corresponding parameters in the presence of 14% glycerol were L(pg)[cpa]=1.063 x 10(-15)m3/Ns (0.0063 microm/min atm), and E(Lp)[cpa]=83.81 kJ/mol (20.04 kcal/mol) (R2=0.997). The parameters in the presence of 10% DMSO were L(pg)[cpa]=1.4 x 10(-15)m3/Ns (0.0083 microm/min atm), and E(Lp)[cpa]=90.96 kJ/mol (21.75 kcal/mol) (R2=0.996). Parameters obtained in this study suggested that the optimal rate of cooling for X. helleri sperm cells in the presence of CPAs ranged from 20 to 35 degrees C/min and were in close agreement with recently published, empirically determined optimal cooling rates.
Figures
Similar articles
-
Subzero water permeability parameters and optimal freezing rates for sperm cells of the southern platyfish, Xiphophorus maculatus.Cryobiology. 2005 Jun;50(3):250-63. doi: 10.1016/j.cryobiol.2005.02.003. Cryobiology. 2005. PMID: 15925577 Free PMC article.
-
Current status of sperm cryopreservation in biomedical research fish models: zebrafish, medaka, and Xiphophorus.Comp Biochem Physiol C Toxicol Pharmacol. 2009 Mar;149(2):224-32. doi: 10.1016/j.cbpc.2008.07.005. Epub 2008 Jul 18. Comp Biochem Physiol C Toxicol Pharmacol. 2009. PMID: 18691673 Free PMC article. Review.
-
Subzero water permeability parameters of mouse spermatozoa in the presence of extracellular ice and cryoprotective agents.Biol Reprod. 1999 Sep;61(3):764-75. doi: 10.1095/biolreprod61.3.764. Biol Reprod. 1999. PMID: 10456855
-
Cryopreservation of equine sperm: optimal cooling rates in the presence and absence of cryoprotective agents determined using differential scanning calorimetry.Biol Reprod. 2002 Jan;66(1):222-31. doi: 10.1095/biolreprod66.1.222. Biol Reprod. 2002. PMID: 11751286
-
Cryopreservation of boar semen: equilibrium freezing in the cryomicroscope and in straws.Theriogenology. 2005 Jan 15;63(2):383-95. doi: 10.1016/j.theriogenology.2004.09.019. Theriogenology. 2005. PMID: 15626406 Review.
Cited by
-
Sperm vitrification of Prochilodus brevis: influence of diluent, stored volume and supplementation with sulfated polysaccharides of Nile tilapia (Oreochromis niloticus) skin.Anim Reprod. 2025 Feb 7;22(1):e20240075. doi: 10.1590/1984-3143-AR2024-0075. eCollection 2025. Anim Reprod. 2025. PMID: 39935520 Free PMC article.
-
Cryopreservation as a Key Element in the Successful Delivery of Cell-Based Therapies-A Review.Front Med (Lausanne). 2020 Nov 26;7:592242. doi: 10.3389/fmed.2020.592242. eCollection 2020. Front Med (Lausanne). 2020. PMID: 33324662 Free PMC article. Review.
-
Production of F1 interspecies hybrid offspring with cryopreserved sperm from a live-bearing fish, the swordtail Xiphophorus helleri.Biol Reprod. 2007 Mar;76(3):401-6. doi: 10.1095/biolreprod.106.056549. Epub 2006 Nov 15. Biol Reprod. 2007. PMID: 17108333 Free PMC article.
-
Subzero water permeability parameters and optimal freezing rates for sperm cells of the southern platyfish, Xiphophorus maculatus.Cryobiology. 2005 Jun;50(3):250-63. doi: 10.1016/j.cryobiol.2005.02.003. Cryobiology. 2005. PMID: 15925577 Free PMC article.
-
Current status of sperm cryopreservation in biomedical research fish models: zebrafish, medaka, and Xiphophorus.Comp Biochem Physiol C Toxicol Pharmacol. 2009 Mar;149(2):224-32. doi: 10.1016/j.cbpc.2008.07.005. Epub 2008 Jul 18. Comp Biochem Physiol C Toxicol Pharmacol. 2009. PMID: 18691673 Free PMC article. Review.
References
-
- Meyer A. The evolution of sexually selected traits in male swordtail fishes (Xiphophorus:Poeciliidae) Heredity. 1997;79:329–37.
-
- Meyer A, Morrissey JM, Schartl M. Recurrent origin of a sexually selected trait in Xiphophorus fishes inferred from a molecular phylogeny. Nature. 1994;368:539–42. - PubMed
-
- Kallman KD. The sex determining mechanism of the poeciliid fish, Xiphophorus montezumae, and the genetic control of the sexual maturation process and adult size. Copeia. 1983;3:755–69.
-
- Kallman KD. A new look at sex determination in poeciliid fishes. In: Turner BJ, editor. Evolutionary genetics of fishes. New York: Plenum Publishing Corporation; 1984. pp. 95–171.
-
- Tiersch TR, Chandler RW, Kallman KD, Wachtel SS. Estimation of nuclear DNA content by flow cytometry in fishes of the genus Xiphophorus. Comp Biochem Physiol B. 1989;94:465–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
