

Benzodiazepine modulation of partial agonist efficacy and spontaneously active GABA(A) receptors supports an allosteric model of modulation
- PMID: 15912137
- PMCID: PMC1576208
- DOI: 10.1038/sj.bjp.0706251
Benzodiazepine modulation of partial agonist efficacy and spontaneously active GABA(A) receptors supports an allosteric model of modulation
Abstract
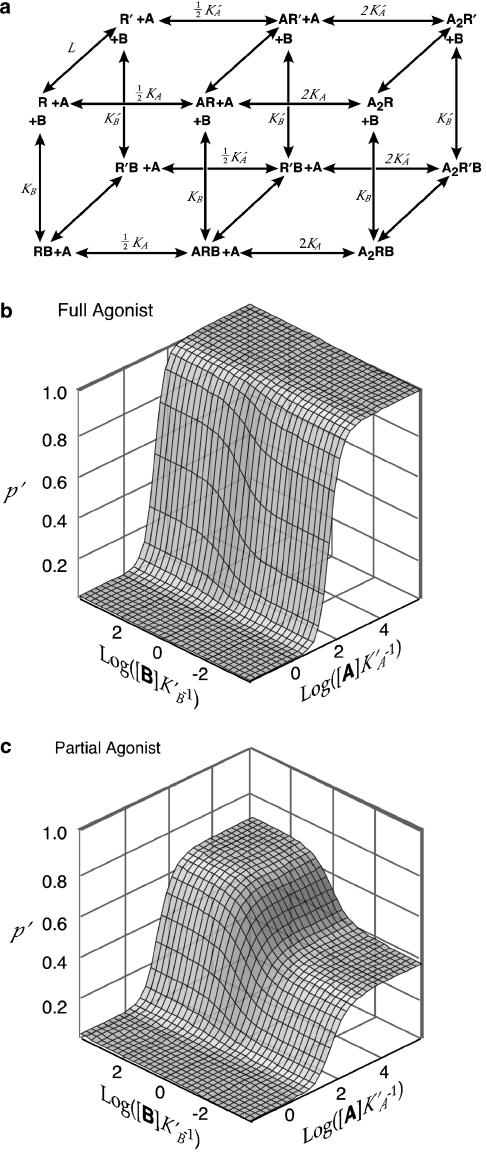
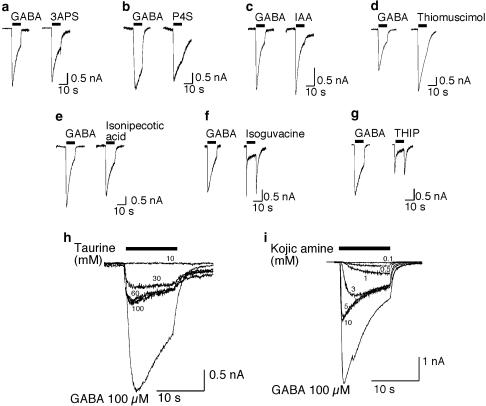
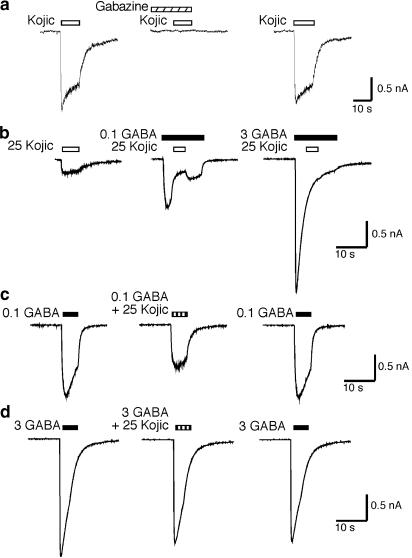
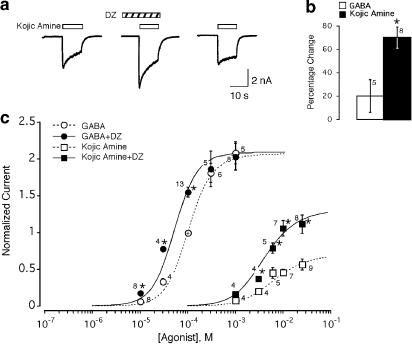
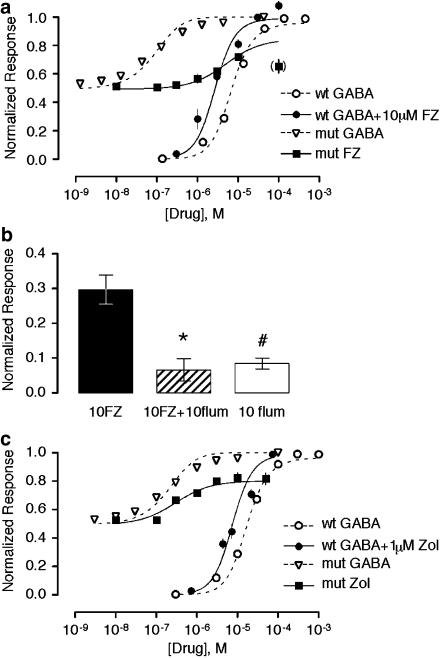
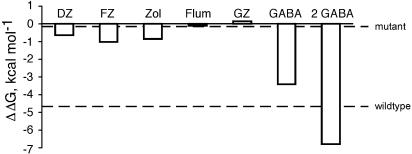
Benzodiazepines (BZDs) have been used extensively for more than 40 years because of their high therapeutic index and low toxicity. Although BZDs are understood to act primarily as allosteric modulators of GABA(A) receptors, the mechanism of modulation is not well understood. The applicability of an allosteric model with two binding sites for gamma-aminobutyric acid (GABA) and one for a BZD-like modulator was investigated. This model predicts that BZDs should enhance the efficacy of partial agonists. Consistent with this prediction, diazepam increased the efficacy of the GABA(A) receptor partial agonist kojic amine in chick spinal cord neurons. To further test the validity of the model, the effects of diazepam, flurazepam, and zolpidem were examined using wild-type and spontaneously active mutant alpha1(L263S)beta3gamma2 GABA(A) receptors expressed in HEK-293 cells. In agreement with the predictions of the allosteric model, all three modulators acted as direct agonists for the spontaneously active receptors. The results indicate that BZD-like modulators enhance the amplitude of the GABA response by stabilizing the open channel active state relative to the inactive state by less than 1 kcal, which is similar to the energy of stabilization conferred by a single hydrogen bond.
Figures
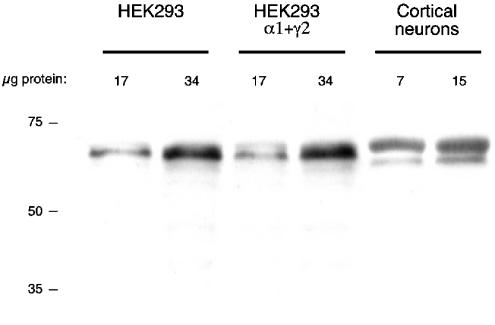
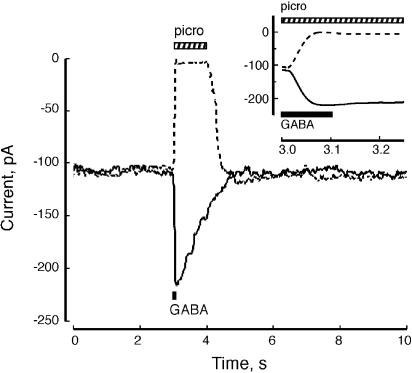
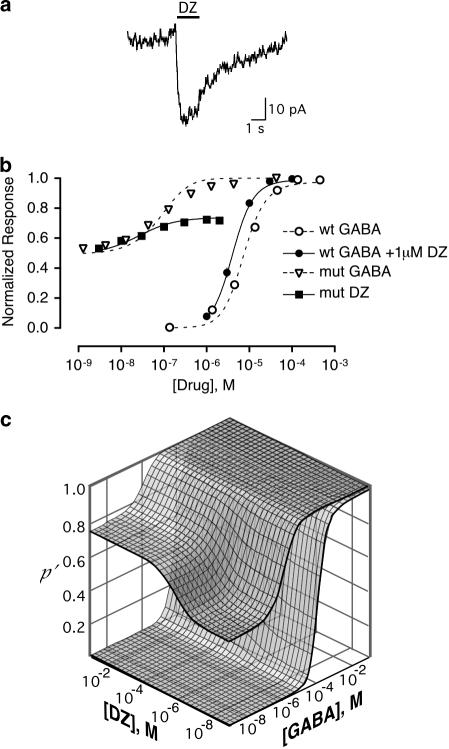
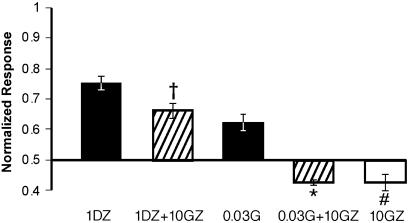
References
-
- CELENTANO J.J., GIBBS T.T., FARB D.H. Negative modulation of the GABA response by extracellular zinc. Mol. Pharmacol. 1991;40:766–773. - PubMed
-
- CHOI D.W., FARB D.H., FISCHBACH G.D. Chlordiazepoxide selectively augments GABA action in spinal cord cell cultures. Nat. (London) 1977;269:342–344. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
