

Cone-like morphological, molecular, and electrophysiological features of the photoreceptors of the Nrl knockout mouse
- PMID: 15914637
- PMCID: PMC4505833
- DOI: 10.1167/iovs.04-1427
Cone-like morphological, molecular, and electrophysiological features of the photoreceptors of the Nrl knockout mouse
Abstract
Purpose: To test the hypothesis that Nrl(-)(/)(-) photoreceptors are cones, by comparing them with WT rods and cones using morphological, molecular, histochemical, and electrophysiological criteria.
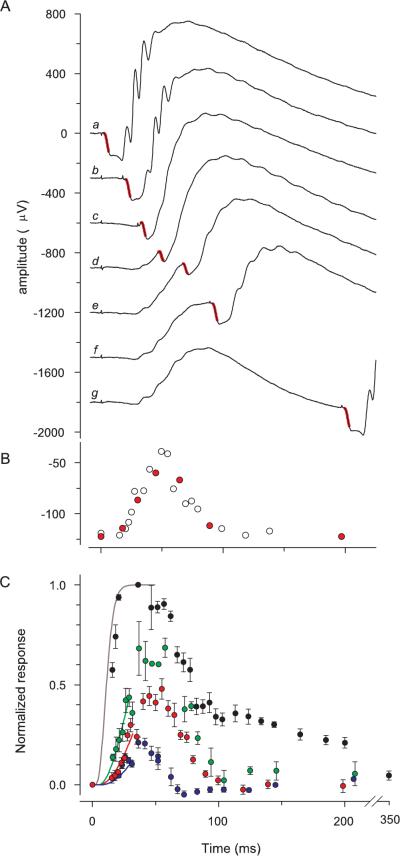
Methods: The photoreceptor layer of fixed retinal tissue of 4- to 6-week-old mice was examined in plastic sections by electron microscopy, and by confocal microscopy in frozen sections immunolabeled for the mouse UV-cone pigment and colabeled with PNA. Quantitative immunoblot analysis was used to determine the levels of expression of key cone-specific proteins. Single- and paired-flash methods were used to extract the spectral sensitivity, kinetics, and amplification of the a-wave of the ERG.
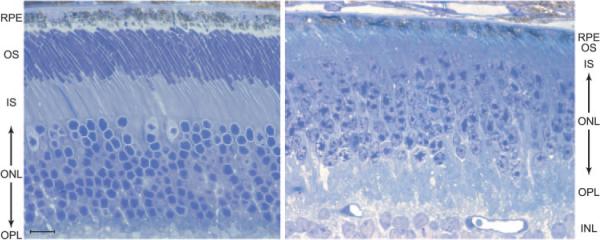
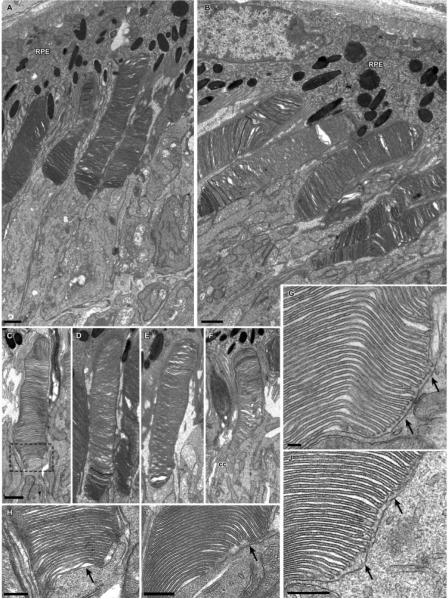
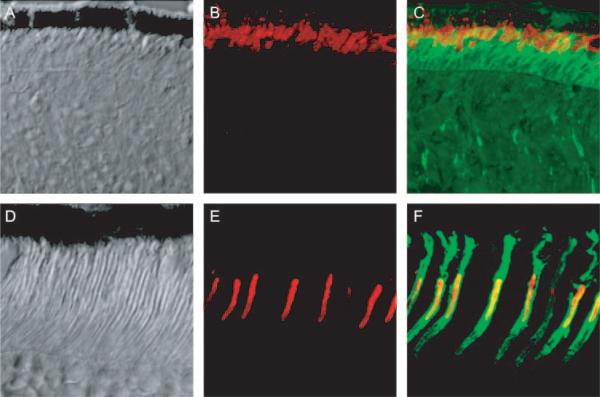
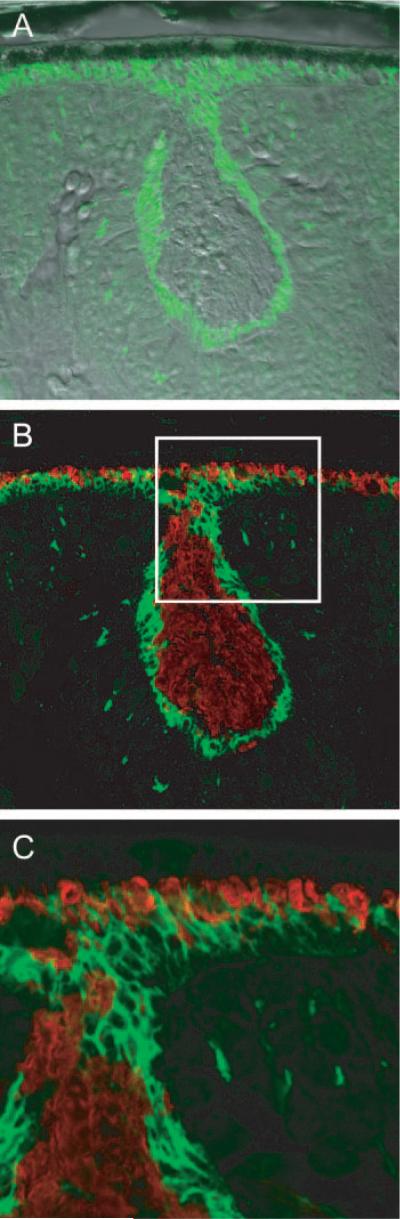
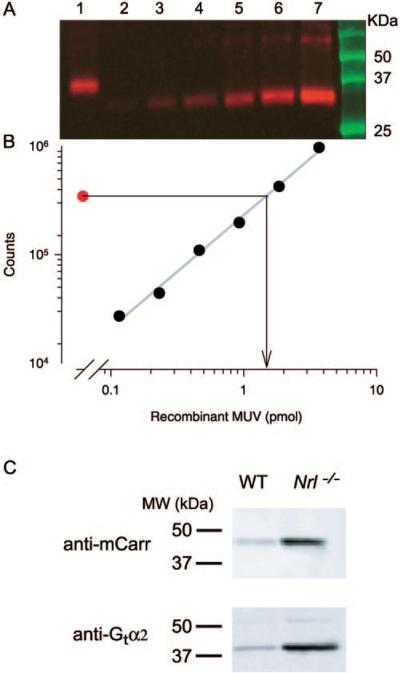
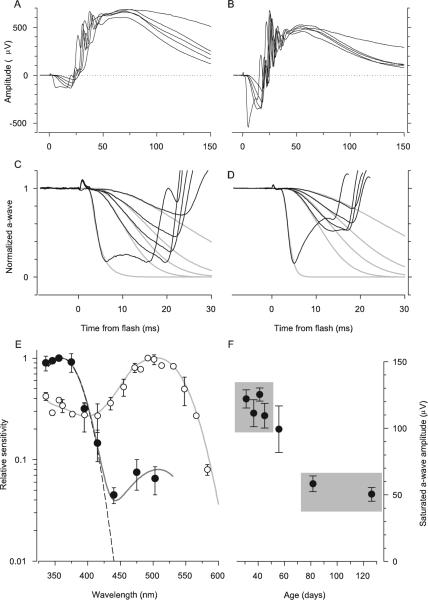
Results: Outer segments of Nrl(-/-) photoreceptors ( approximately 7 mum) are shorter than those of wild-type (WT) rods ( approximately 25 mum) and cones ( approximately 15 mum); but, like WT cones, they have 25 or more basal discs open to the extracellular space, extracellular matrix sheaths stained by PNA, chromatin "clumping" in their nuclei, and mitochondria two times shorter than rods. Nrl(-/-) photoreceptors express the mouse UV cone pigment, cone transducin, and cone arrestin in amounts expected, given the relative size and density of cones in the two retinas. The ERG a-wave was used to assay the properties of the photocurrent response. The sensitivity of the Nrl(-/-) a-wave is at its maximum at 360 nm, with a secondary mode at 510 nm having approximately one-tenth the maximum sensitivity. These wavelengths are the lambda(max) of the two mouse cone pigments. The time to peak of the dim-flash photocurrent response was approximately 50 ms, more than two times faster than that of rods.
Conclusions: Many morphological, molecular, and electrophysiological features of the Nrl(-/-) photoreceptors are cone-like, and strongly distinguish these cells from rods. This retina provides a model for the investigation of cone function and cone-specific genetic disease.
Figures
References
-
- Swain PK, Hicks D, Mears AJ, et al. Multiple phosphorylated iso-forms of NRL are expressed in rod photoreceptors. J Biol Chem. 2001;276:36824–36830. - PubMed
-
- Yoshida S, Mears AJ, Friedman JS, et al. Expression profiling of the developing and mature Nrl−/− mouse retina: identification of retinal disease candidates and transcriptional regulatory targets of Nrl. Hum Mol Genet. 2004;13:1487–1503. - PubMed
-
- Bessant DA, Payne AM, Mitton KP, et al. A mutation in NRL is associated with autosomal dominant retinitis pigmentosa. Nat Genet. 1999;21:355–356. - PubMed
-
- Mears AJ, Kondo M, Swain PK, et al. Nrl is required for rod photoreceptor development. Nat Genet. 2001;29:447–452. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
