

The peroxisome proliferator-activated receptor-gamma regulates murine pyruvate carboxylase gene expression in vivo and in vitro
- PMID: 15917242
- PMCID: PMC4304003
- DOI: 10.1074/jbc.M503836200
The peroxisome proliferator-activated receptor-gamma regulates murine pyruvate carboxylase gene expression in vivo and in vitro
Abstract
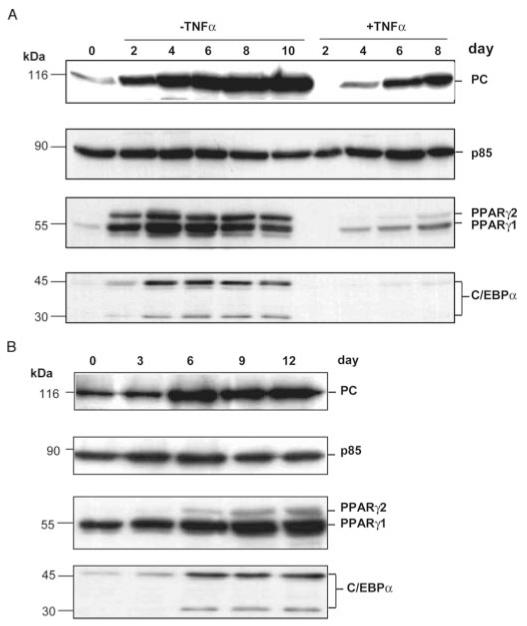
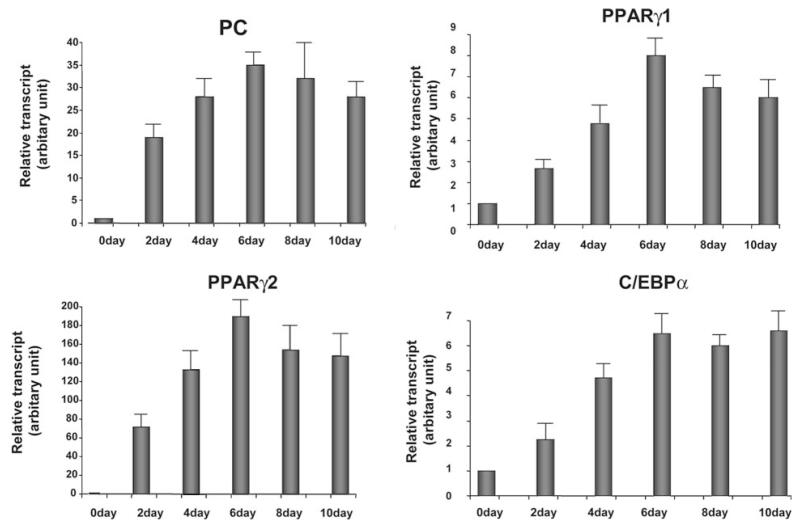
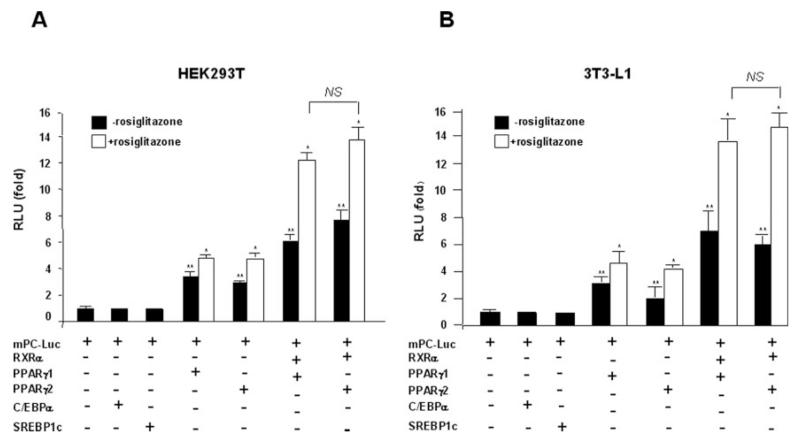
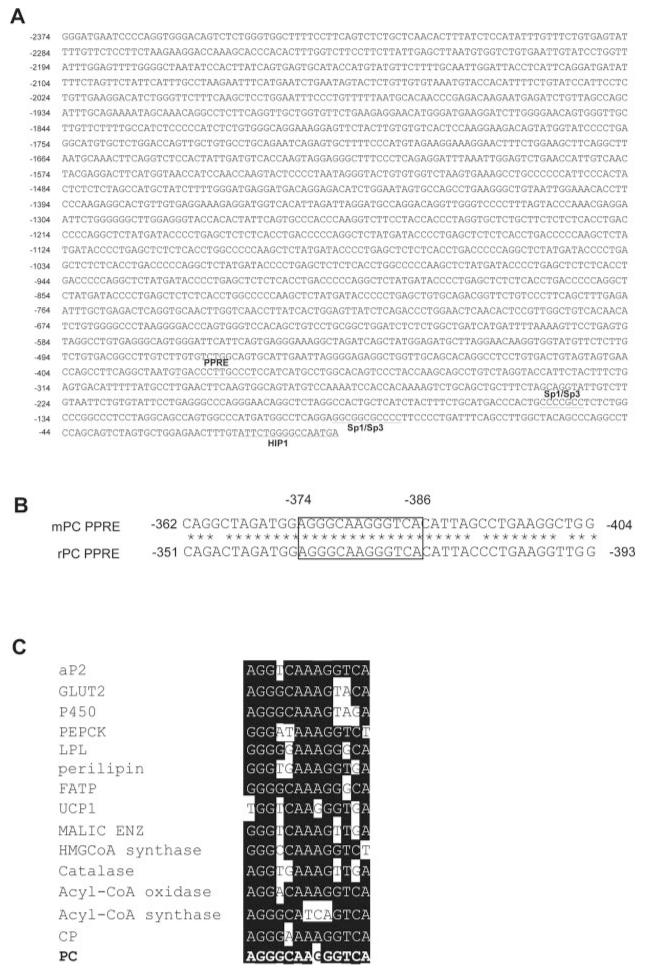
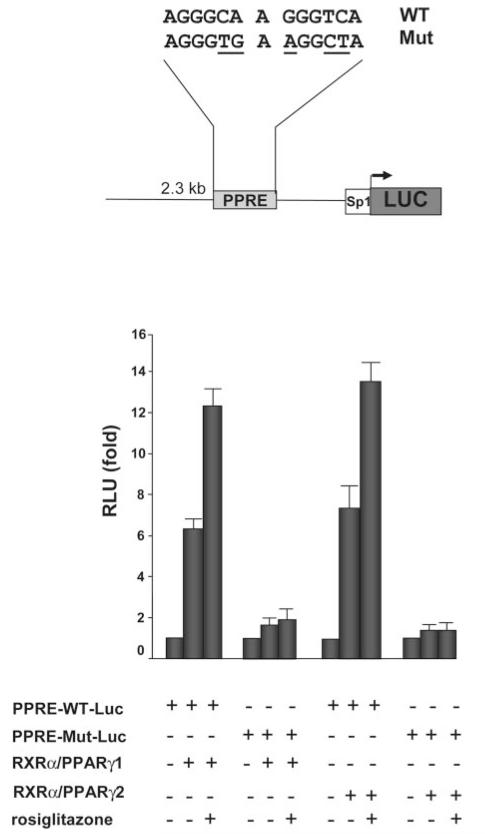
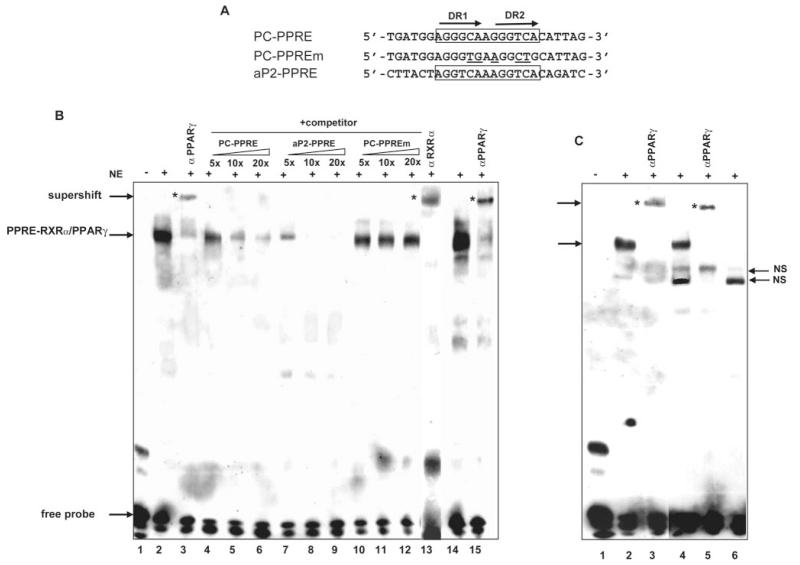
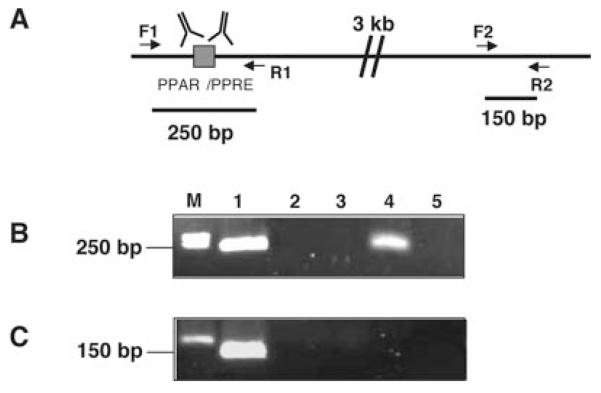
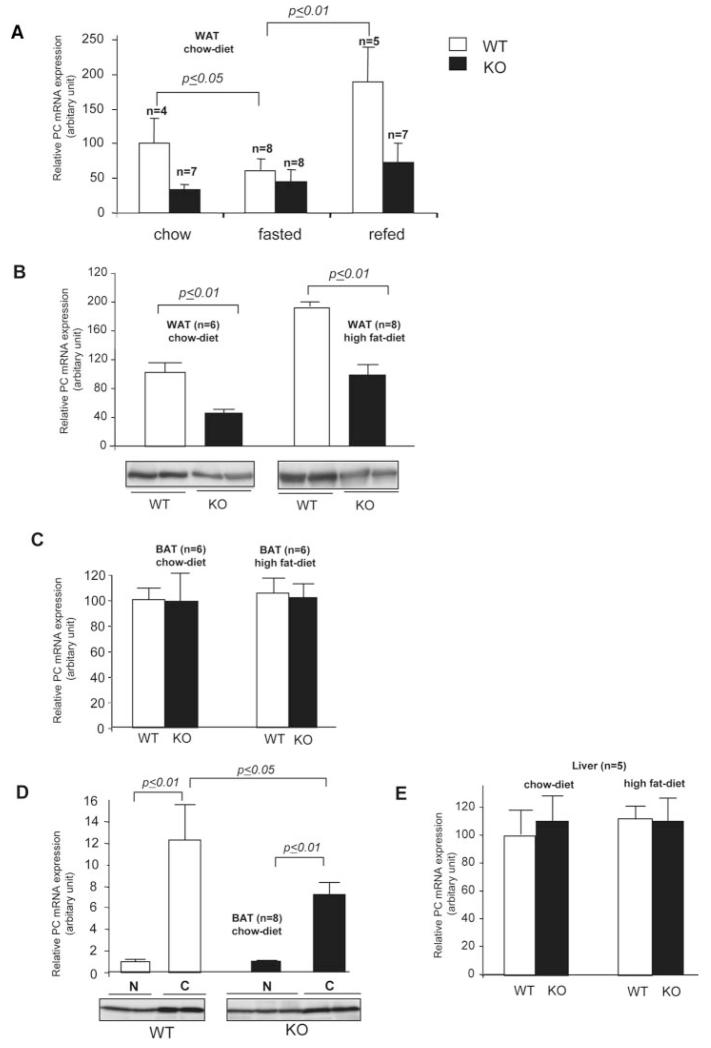
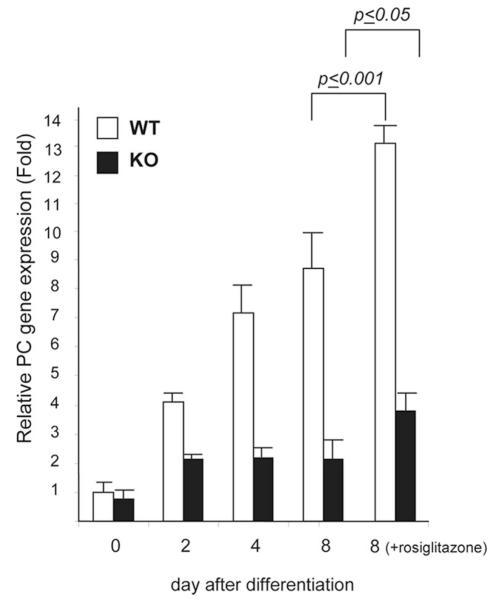
Pyruvate carboxylase (PC) plays a crucial role in various metabolic pathways, including gluconeogenesis, lipogenesis, and glucose-induced insulin secretion. Here we showed for the first time that the PC gene is transcriptionally regulated by peroxisome proliferator-activated receptor-gamma (PPARgamma) in vitro and in vivo in white and brown adipose tissue. PC mRNA and protein are markedly increased during differentiation of 3T3-L1 cells and HIB-1B, in parallel with the expression of the adipogenic transcription factors, CCAAT-enhancer binding protein alpha, PPARgamma1, and PPARgamma2. Tumor necrosis factor-alpha, a cytokine that blocks differentiation of 3T3-L1 cells, suppressed PC expression. Co-transfection studies in 3T3-L1 preadipocytes or HEK293T cells with a 2.3-kb promoter fragment of mouse PC gene linked to a luciferase reporter construct and with plasmids overexpressing retinoid X receptor alpha/PPARgamma1 or retinoid X receptor alpha/PPARgamma2 showed a 6-8-fold increase above the basal promoter activity. Furthermore, treatment of these transfected cells with the PPARgamma agonist doubled the promoter activity. Mutation of the putative PPAR-response element-(-386/-374) of this 2.3-kb PC promoter fragment abolished the PPARgamma response. Gel shift and chromatin immunoprecipitation assays demonstrated that endogenous PPARgamma binds to this functional PPAR-response element of the PC promoter. Mice with targeted disruption of the PPARgamma2 gene displayed approximately 50-60% reduction of PC mRNA and protein in white adipose tissue. Similarly, in brown adipose tissue of PPARgamma2-deficient mice subjected to cold exposure, PC mRNA was 40% lower than that of wild type mice. Impaired in vitro differentiation of white adipocytes of PPARgamma2 knock-out mice was also associated with a marked reduction of PC mRNA. Our findings identified PC as a PPARgamma-regulated gene and suggested a role for PPARgamma regulating intermediary metabolism.
Figures
References
-
- Mackall JC, Student AK, Polakis SE, Lane MD. J. Biol. Chem. 1976;251:6462–6464. - PubMed
-
- Cowherd RM, Lyle RE, McGehee RE., Jr. Semin. Cell. Dev. Biol. 1999;10:3–10. - PubMed
-
- Rosen ED, Walkey CJ, Puigserver P, Spiegelman BM. Genes Dev. 2000;14:1293–1307. - PubMed
-
- Rosen ED, Spiegelman BM. Annu. Rev. Cell Dev. Biol. 2000;16:145–171. - PubMed
-
- Rosen ED, Spiegelman BM. J. Biol. Chem. 2001;276:37731–37734. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
