

Intestinal dysplasia induced by simian virus 40 T antigen is independent of p53
- PMID: 15919904
- PMCID: PMC1143657
- DOI: 10.1128/JVI.79.12.7492-7502.2005
Intestinal dysplasia induced by simian virus 40 T antigen is independent of p53
Abstract
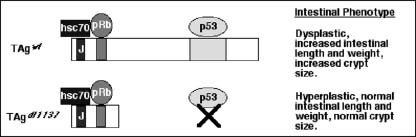
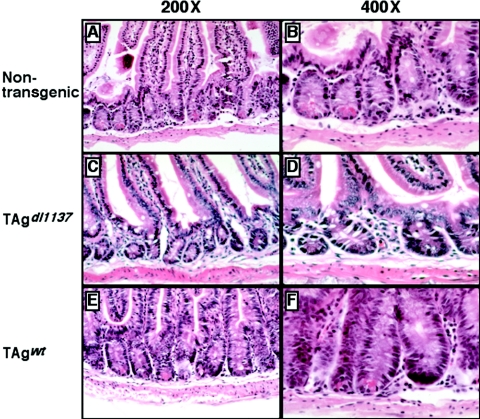
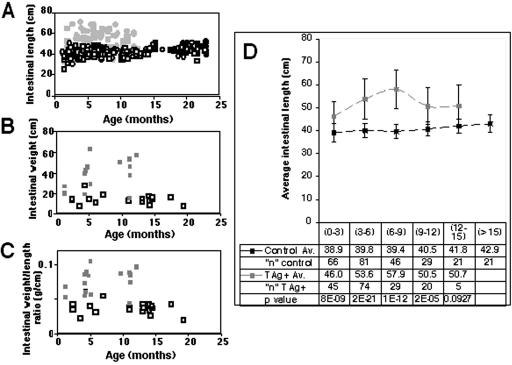
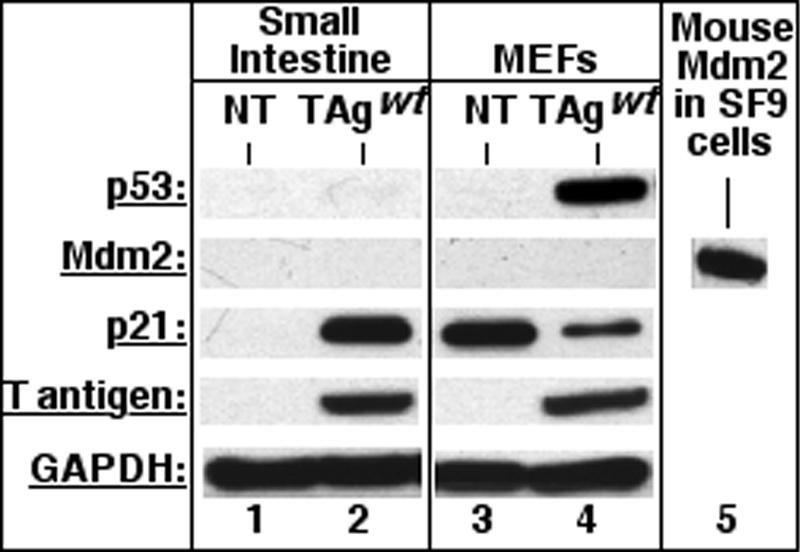
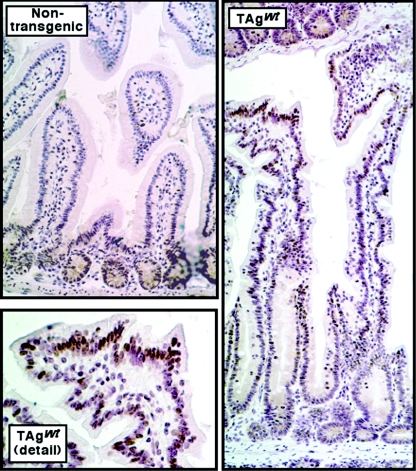
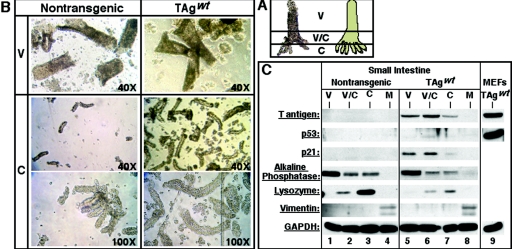
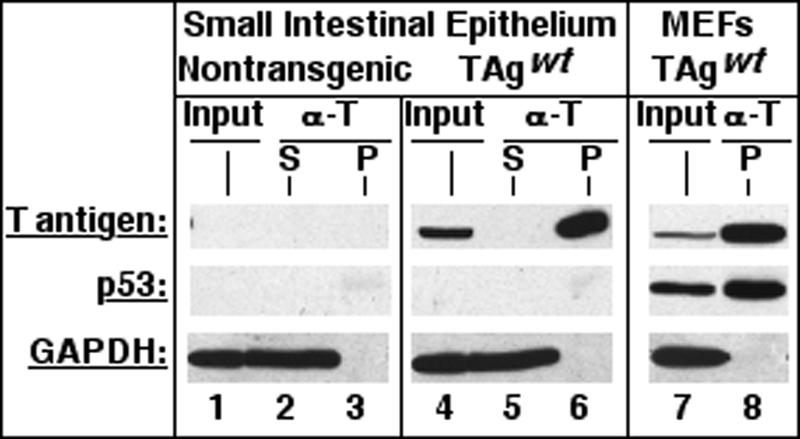
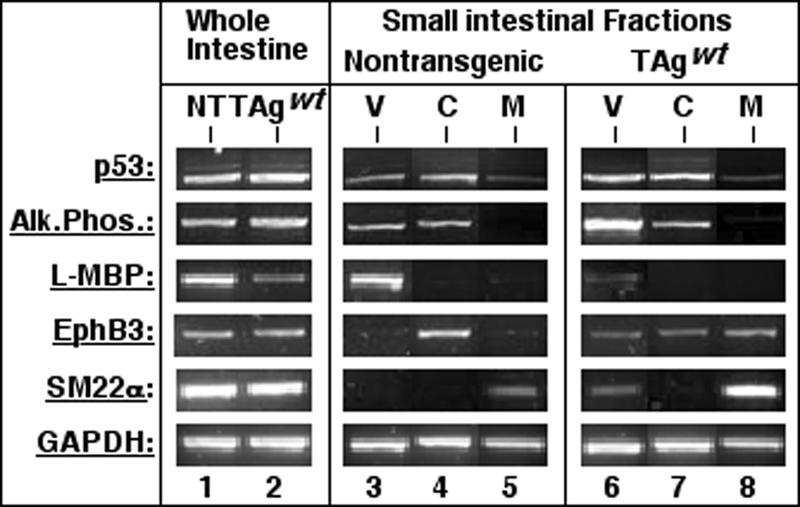
Transgenic mice expressing simian virus 40 large T antigen in enterocytes develop intestinal hyperplasia that progresses to dysplasia with age. Hyperplasia is dependent on T antigen binding to the retinoblastoma (pRb) family of tumor suppressor proteins. Mice expressing a truncated T antigen that inactivates the pRb-family, but is defective for binding p53, exhibit hyperplasia but do not progress to dysplasia. We hypothesized that the inhibition of the pRb family leads to entry of enterocytes into the cell cycle, resulting in hyperplasia, while inactivation of p53 is required for progression to dysplasia. Therefore, we examined T antigen/p53 complexes from the intestines of transgenic mice. We found that T antigen did not induce p53 stabilization, and we could not detect T antigen/p53 complexes in villus enterocytes. In contrast, T antigen expression led to a large increase in the levels of the cyclin-dependent kinase inhibitor p21. Furthermore, mice in which pRb was inactivated by a truncated T antigen in a p53 null background exhibited intestinal hyperplasia but no progression to dysplasia. These data indicate that loss of p53 function does not play a role in T antigen-induced dysplasia in the intestine. Rather, some unknown function of T antigen is essential for progression beyond hyperplasia.
Figures
Similar articles
-
Enterocyte proliferation and intestinal hyperplasia induced by simian virus 40 T antigen require a functional J domain.J Virol. 2007 Sep;81(17):9481-9. doi: 10.1128/JVI.00922-07. Epub 2007 Jun 20. J Virol. 2007. PMID: 17581980 Free PMC article.
-
Intestinal hyperplasia induced by simian virus 40 large tumor antigen requires E2F2.J Virol. 2007 Dec;81(23):13191-9. doi: 10.1128/JVI.01658-07. Epub 2007 Sep 12. J Virol. 2007. PMID: 17855529 Free PMC article.
-
Simian virus 40 T-antigen-mediated gene regulation in enterocytes is controlled primarily by the Rb-E2F pathway.J Virol. 2009 Sep;83(18):9521-31. doi: 10.1128/JVI.00583-09. Epub 2009 Jul 1. J Virol. 2009. PMID: 19570859 Free PMC article.
-
Induction versus progression of brain tumor development: differential functions for the pRB- and p53-targeting domains of simian virus 40 T antigen.Mol Cell Biol. 1994 Apr;14(4):2686-98. doi: 10.1128/mcb.14.4.2686-2698.1994. Mol Cell Biol. 1994. PMID: 8139568 Free PMC article.
-
Interactions between SV40 large-tumor antigen and the growth suppressor proteins pRB and p53.FASEB J. 1993 Jul;7(10):866-71. doi: 10.1096/fasebj.7.10.8344486. FASEB J. 1993. PMID: 8344486 Review.
Cited by
-
Polyomavirus T antigens activate an antiviral state.Virology. 2015 Feb;476:377-385. doi: 10.1016/j.virol.2014.12.032. Epub 2015 Jan 10. Virology. 2015. PMID: 25589241 Free PMC article.
-
Enterocyte proliferation and intestinal hyperplasia induced by simian virus 40 T antigen require a functional J domain.J Virol. 2007 Sep;81(17):9481-9. doi: 10.1128/JVI.00922-07. Epub 2007 Jun 20. J Virol. 2007. PMID: 17581980 Free PMC article.
-
A retinoblastoma allele that is mutated at its common E2F interaction site inhibits cell proliferation in gene-targeted mice.Mol Cell Biol. 2014 Jun;34(11):2029-45. doi: 10.1128/MCB.01589-13. Epub 2014 Mar 24. Mol Cell Biol. 2014. PMID: 24662053 Free PMC article.
-
The retinoblastoma tumor suppressor regulates a xenobiotic detoxification pathway.PLoS One. 2011;6(10):e26019. doi: 10.1371/journal.pone.0026019. Epub 2011 Oct 12. PLoS One. 2011. PMID: 22022495 Free PMC article.
-
Prevention of senescence progression in reversibly immortalized human ensheathing glia permits their survival after deimmortalization.Mol Ther. 2010 Feb;18(2):394-403. doi: 10.1038/mt.2009.268. Epub 2009 Nov 24. Mol Ther. 2010. PMID: 19935779 Free PMC article.
References
-
- Bargonetti, J., I. Reynisdottir, P. N. Friedman, and C. Prives. 1992. Site-specific binding of wild-type p53 to cellular DNA is inhibited by SV40 T antigen and mutant p53. Genes Dev. 6:1886-1898. - PubMed
-
- Batlle, E., J. T. Henderson, H. Beghtel, M. M. W. van den Born, E. Sancho, G. Huls, J. Meeldijk, J. Robertson, M. van de Wetering, T. Pawson, and H. Clevers. 2002. β-Catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/EphrinB. Cell 111:251-263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
