

Identification of the rabies virus alpha/beta interferon antagonist: phosphoprotein P interferes with phosphorylation of interferon regulatory factor 3
- PMID: 15919920
- PMCID: PMC1143667
- DOI: 10.1128/JVI.79.12.7673-7681.2005
Identification of the rabies virus alpha/beta interferon antagonist: phosphoprotein P interferes with phosphorylation of interferon regulatory factor 3
Abstract
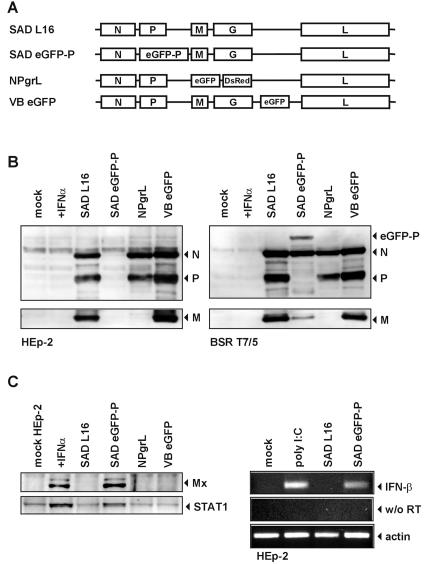
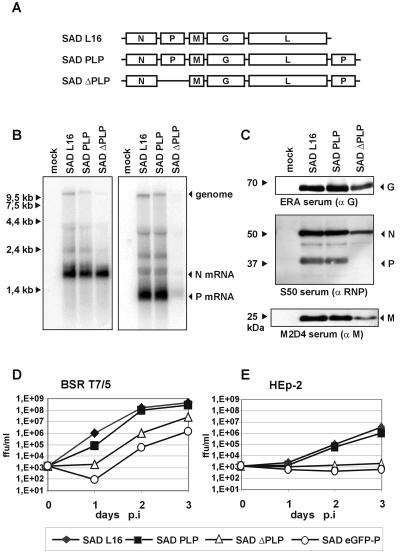
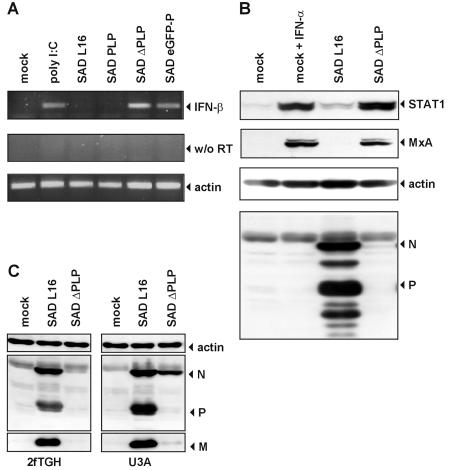
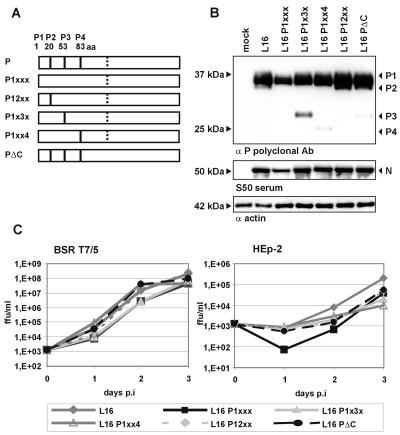
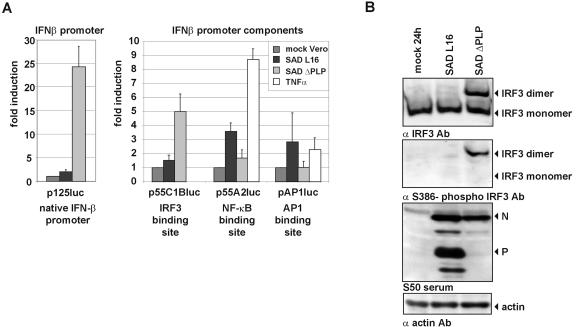
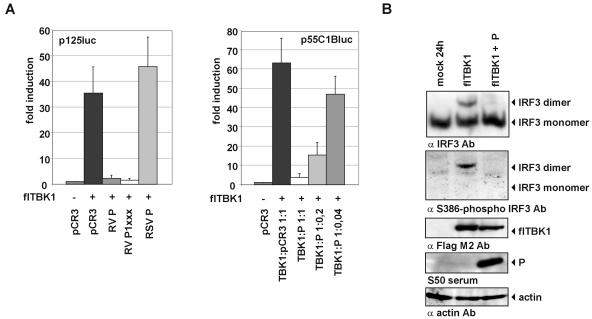
Rabies virus (RV) of the Rhabdoviridae family grows in alpha/beta interferon (IFN)-competent cells, suggesting the existence of viral mechanisms preventing IFN gene expression. We here identify the viral phosphoprotein P as the responsible IFN antagonist. The critical involvement of P was first suggested by the observation that an RV expressing an enhanced green fluorescent protein (eGFP)-P fusion protein (SAD eGFP-P) (S. Finke, K. Brzozka, and K. K. Conzelmann, J. Virol. 78:12333-12343, 2004) was eliminated in IFN-competent HEp-2 cell cultures, in contrast to wild-type (wt) RV or an RV replicon lacking the genes for matrix protein and glycoprotein. SAD eGFP-P induced transcription of the IFN-beta gene and expression of the IFN-responsive MxA and STAT-1 genes. Similarly, an RV expressing low levels of P, which was generated by moving the P gene to a promoter-distal gene position (SAD DeltaPLP), lost the ability to prevent IFN induction. The analysis of RV mutants lacking expression of truncated P proteins P2, P3, or P4, which are expressed from internal AUG codons of the wt RV P open reading frame, further showed that full-length P is competent in suppressing IFN-beta gene expression. In contrast to wt RV, the IFN-inducing SAD DeltaPLP caused S386 phosphorylation, dimerization, and transcriptional activity of IFN regulatory factor 3 (IRF-3). Phosphorylation of IRF-3 by TANK-binding kinase-1 expressed from transfected plasmids was abolished in wt RV-infected cells or by cotransfection of P-encoding plasmids. Thus, RV P is necessary and sufficient to prevent a critical IFN response in virus-infected cells by targeting activation of IRF-3 by an upstream kinase.
Figures
References
-
- Ahmed, M., M. O. McKenzie, S. Puckett, M. Hojnacki, L. Poliquin, and D. S. Lyles. 2003. Ability of the matrix protein of vesicular stomatitis virus to suppress beta interferon gene expression is genetically correlated with the inhibition of host RNA and protein synthesis. J. Virol. 77:4646-4657. - PMC - PubMed
-
- Akira, S., and K. Takeda. 2004. Toll-like receptor signalling. Nat. Rev. Immunol. 4:499-511. - PubMed
-
- Basler, C. F., and A. Garcia-Sastre. 2002. Viruses and the type I interferon antiviral system: induction and evasion. Int. Rev. Immunol. 21:305-337. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
