

Allosteric effects of ligands and mutations on poliovirus RNA-dependent RNA polymerase
- PMID: 15919933
- PMCID: PMC1143668
- DOI: 10.1128/JVI.79.12.7803-7811.2005
Allosteric effects of ligands and mutations on poliovirus RNA-dependent RNA polymerase
Abstract
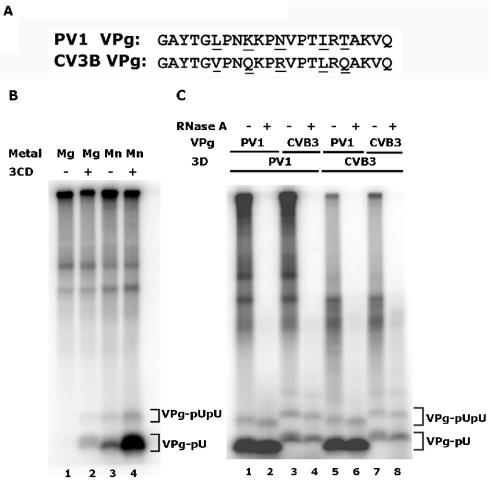
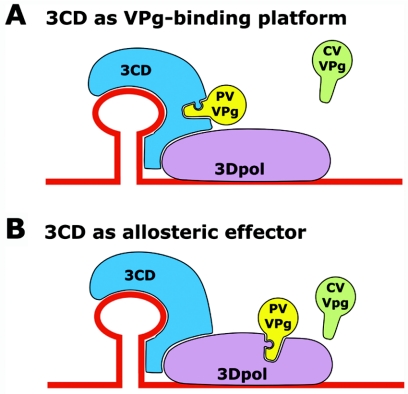
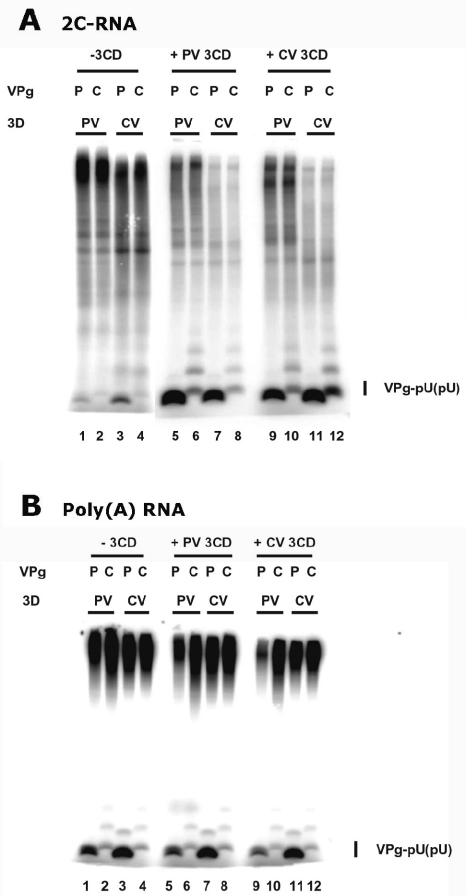
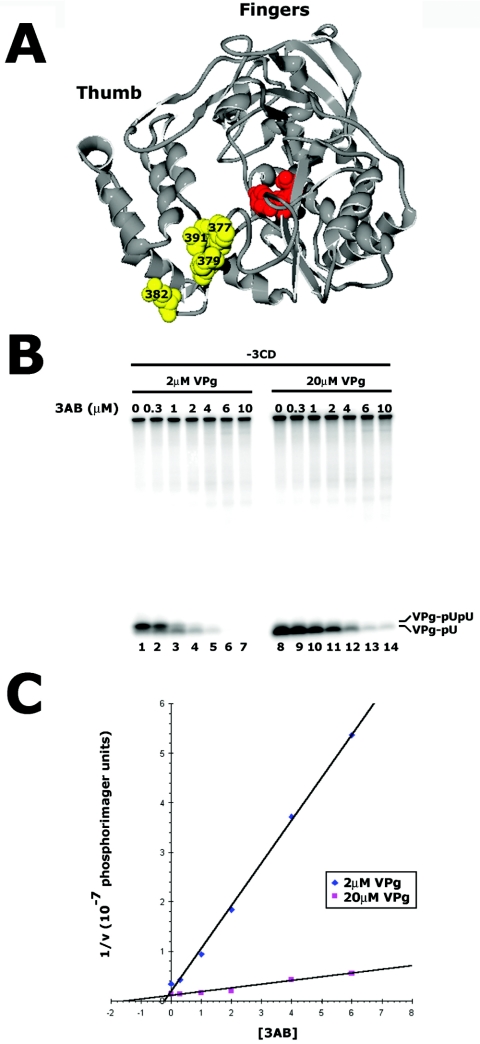
Protein priming of viral RNA synthesis plays an essential role in the replication of picornavirus RNA. Both poliovirus and coxsackievirus encode a small polypeptide, VPg, which serves as a primer for addition of the first nucleotide during synthesis of both positive and negative strands. This study examined the effects on the VPg uridylylation reaction of the RNA template sequence, the origin of VPg (coxsackievirus or poliovirus), the origin of 3D polymerase (coxsackievirus or poliovirus), the presence and origin of interacting protein 3CD, and the introduction of mutations at specific regions in the poliovirus 3D polymerase. Substantial effects associated with VPg origin were traced to differences in VPg-polymerase interactions. The effects of 3CD proteins and mutations at polymerase-polymerase intermolecular Interface I were most consistent with allosteric effects on the catalytic 3D polymerase molecule. In conclusion, the efficiency and specificity of VPg uridylylation by picornavirus polymerases is greatly influenced by allosteric effects of ligand binding that are likely to be relevant during the viral replicative cycle.
Figures
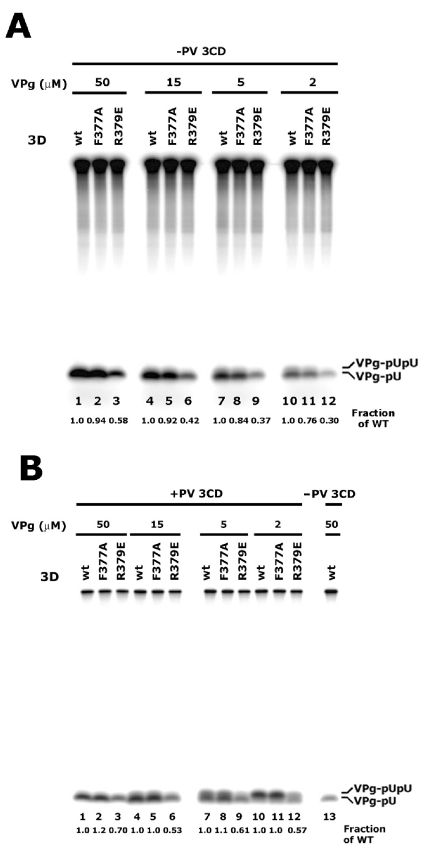
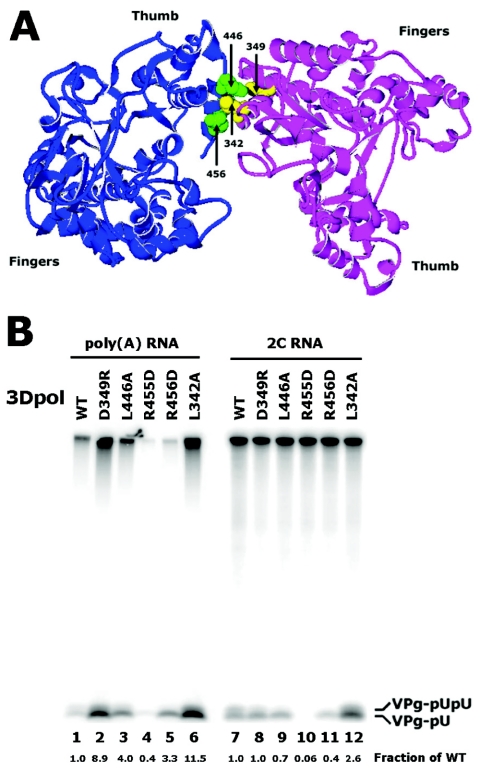
References
-
- Cheetham, G. M. T., and T. A. Steitz. 1999. Structure of a transcribing T7 RNA polymerase initiation complex. Science 286:2305-2309. - PubMed
-
- Crowder, S., and K. Kirkegaard. 2004. Complete three-dimensional structures of picornaviral RNA-dependent RNA polymerases. Structure 12:1336-1339. - PubMed
-
- Dayhuff, T. J., R. F. Gesteland, and J. F. Atkins. 1992. Electrophoresis, autoradiography and electroblotting of peptides: T4 gene 60 hopping. BioTechniques 13:500-503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
