

Stringent requirement for the C protein of wild-type measles virus for growth both in vitro and in macaques
- PMID: 15919937
- PMCID: PMC1143652
- DOI: 10.1128/JVI.79.12.7838-7844.2005
Stringent requirement for the C protein of wild-type measles virus for growth both in vitro and in macaques
Abstract
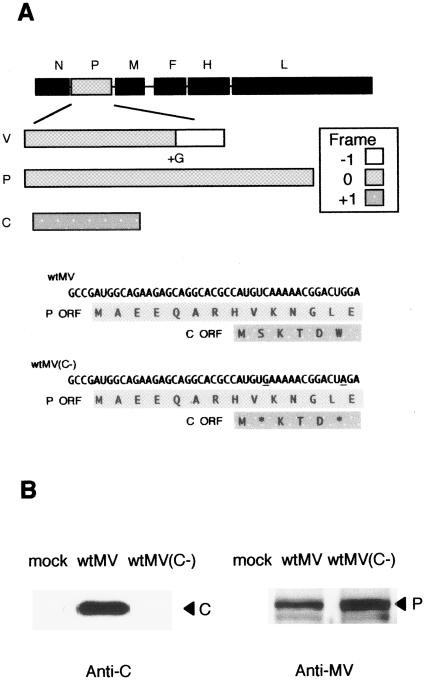
The P gene of measles virus (MV) encodes the P protein and three accessory proteins (C, V, and R). However, the role of these accessory proteins in the natural course of MV infection remains unclear. For this study, we generated a recombinant wild-type MV lacking the C protein, called wtMV(C-), by using a reverse genetics system (M. Takeda, K. Takeuchi, N. Miyajima, F. Kobune, Y. Ami, N. Nagata, Y. Suzaki, Y. Nagai, and M. Tashiro, J. Virol. 74:6643-6647). When 293 cells expressing the MV receptor SLAM (293/hSLAM) were infected with wtMV(C-) or parental wild-type MV (wtMV), the growth of wtMV(C-) was restricted, particularly during late stages. Enhanced green fluorescent protein-expressing wtMV(C-) consistently induced late-stage cell rounding and cell death in the presence of a fusion-inhibiting peptide, suggesting that the C protein can prevent cell death and is required for long-term MV infection. Neutralizing antibodies against alpha/beta interferon did not restore the growth restriction of wtMV(C-) in 293/hSLAM cells. When cynomolgus monkeys were infected with wtMV(C-) or wtMV, the number of MV-infected cells in the thymus was >1,000-fold smaller for wtMV(C-) than for wtMV. Immunohistochemical analyses showed strong expression of an MV antigen in the spleen, lymph nodes, tonsils, and larynx of a cynomolgus monkey infected with wtMV but dramatically reduced expression in the same tissues in a cynomolgus monkey infected with wtMV(C-). These data indicate that the MV C protein is necessary for efficient MV replication both in vitro and in cynomolgus monkeys.
Figures
Similar articles
-
Wild-type measles virus with the hemagglutinin protein of the edmonston vaccine strain retains wild-type tropism in macaques.J Virol. 2012 Mar;86(6):3027-37. doi: 10.1128/JVI.06517-11. Epub 2012 Jan 11. J Virol. 2012. PMID: 22238320 Free PMC article.
-
V and C proteins of measles virus function as virulence factors in vivo.Virology. 2000 Feb 1;267(1):80-9. doi: 10.1006/viro.1999.0118. Virology. 2000. PMID: 10648185
-
Nectin-4-dependent measles virus spread to the cynomolgus monkey tracheal epithelium: role of infected immune cells infiltrating the lamina propria.J Virol. 2013 Mar;87(5):2526-34. doi: 10.1128/JVI.03037-12. Epub 2012 Dec 19. J Virol. 2013. PMID: 23255790 Free PMC article.
-
Measles virus attenuation associated with transcriptional impediment and a few amino acid changes in the polymerase and accessory proteins.J Virol. 1998 Nov;72(11):8690-6. doi: 10.1128/JVI.72.11.8690-8696.1998. J Virol. 1998. PMID: 9765410 Free PMC article.
-
Nectin 4 is the epithelial cell receptor for measles virus.Trends Microbiol. 2012 Sep;20(9):429-39. doi: 10.1016/j.tim.2012.05.006. Epub 2012 Jun 20. Trends Microbiol. 2012. PMID: 22721863 Review.
Cited by
-
Wild-Type Measles Virus is Intrinsically Dual-Tropic.Front Microbiol. 2012 Jan 13;2:279. doi: 10.3389/fmicb.2011.00279. eCollection 2011. Front Microbiol. 2012. PMID: 22347873 Free PMC article.
-
Peste des petits ruminants virus non-structural C protein inhibits the induction of interferon-β by potentially interacting with MAVS and RIG-I.Virus Genes. 2021 Feb;57(1):60-71. doi: 10.1007/s11262-020-01811-y. Epub 2021 Jan 3. Virus Genes. 2021. PMID: 33389635 Free PMC article.
-
Measles virus V protein is a decoy substrate for IkappaB kinase alpha and prevents Toll-like receptor 7/9-mediated interferon induction.J Virol. 2008 Dec;82(24):12365-73. doi: 10.1128/JVI.01321-08. Epub 2008 Oct 15. J Virol. 2008. PMID: 18922877 Free PMC article.
-
Cell tropism and pathogenesis of measles virus in monkeys.Front Microbiol. 2012 Jan 30;3:14. doi: 10.3389/fmicb.2012.00014. eCollection 2012. Front Microbiol. 2012. PMID: 22363320 Free PMC article.
-
Paramyxovirus disruption of interferon signal transduction: STATus report.J Interferon Cytokine Res. 2009 Sep;29(9):531-7. doi: 10.1089/jir.2009.0070. J Interferon Cytokine Res. 2009. PMID: 19694544 Free PMC article. Review.
References
-
- Auwaerter, P. G., P. A. Rota, W. R. Elkins, R. J. Adams, T. DeLozier, Y. Shi, W. J. Bellini, B. R. Murphy, and D. E. Griffin. 1999. Measles virus infection in rhesus macaques: altered immune responses and comparison of the virulence of six different virus strains. J. Infect. Dis. 180:950-958. - PubMed
-
- Curran, J., J. B. Marq, and D. Kolakofsky. 1992. The Sendai virus nonstructural C proteins specifically inhibit viral mRNA synthesis. Virology 189:647-656. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
