

Global analysis of yeast RNA processing identifies new targets of RNase III and uncovers a link between tRNA 5' end processing and tRNA splicing
- PMID: 15920104
- PMCID: PMC1140755
- DOI: 10.1093/nar/gki608
Global analysis of yeast RNA processing identifies new targets of RNase III and uncovers a link between tRNA 5' end processing and tRNA splicing
Abstract
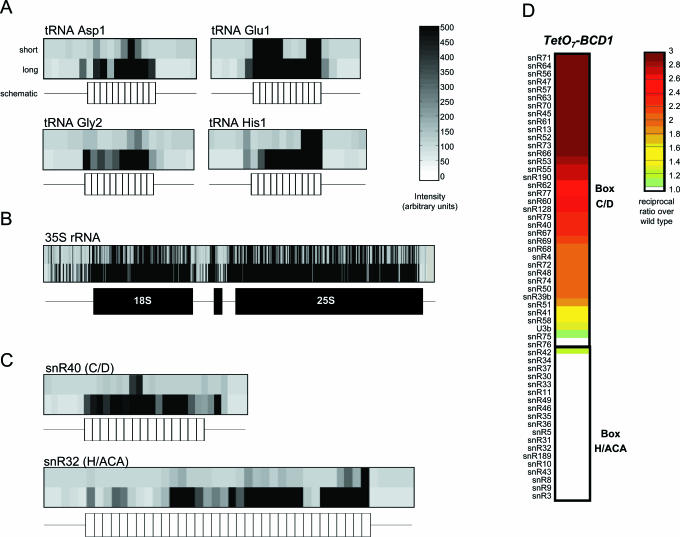
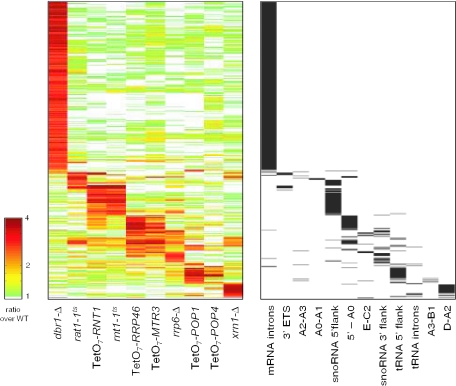
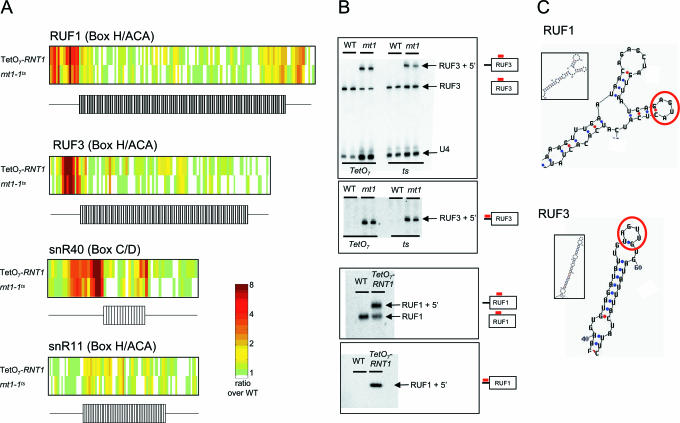
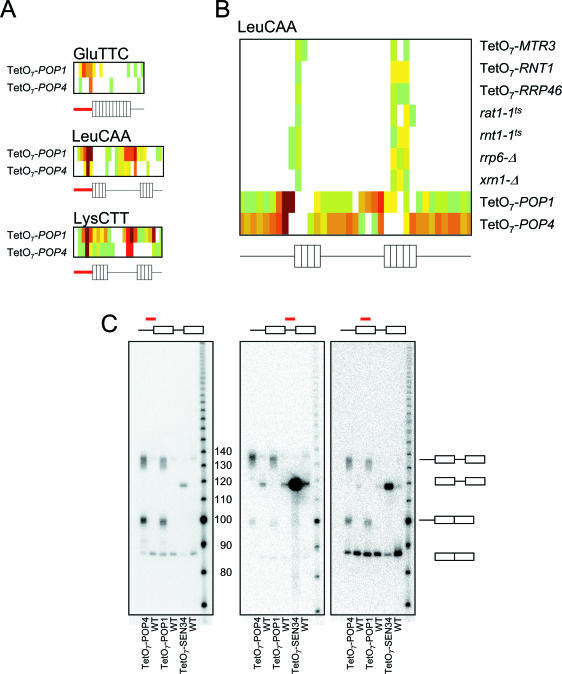
We used a microarray containing probes that tile all known yeast noncoding RNAs (ncRNAs) to investigate RNA biogenesis on a global scale. The microarray verified a general loss of Box C/D snoRNAs in the TetO7-BCD1 mutant, which had previously been shown for only a handful of snoRNAs. We also monitored the accumulation of improperly processed flank sequences of pre-RNAs in strains depleted for known RNA nucleases, including RNase III, Dbr1p, Xrn1p, Rat1p and components of the exosome and RNase P complexes. Among the hundreds of aberrant RNA processing events detected, two novel substrates of Rnt1p (the RUF1 and RUF3 snoRNAs) were identified. We also identified a relationship between tRNA 5' end processing and tRNA splicing, processes that were previously thought to be independent. This analysis demonstrates the applicability of microarray technology to the study of global analysis of ncRNA synthesis and provides an extensive directory of processing events mediated by yeast ncRNA processing enzymes.
Figures
Similar articles
-
Yeast RNase III as a key processing enzyme in small nucleolar RNAs metabolism.J Mol Biol. 1998 Dec 11;284(4):975-88. doi: 10.1006/jmbi.1998.2237. J Mol Biol. 1998. PMID: 9837720
-
Global identification of noncoding RNAs in Saccharomyces cerevisiae by modulating an essential RNA processing pathway.Proc Natl Acad Sci U S A. 2006 Mar 14;103(11):4192-7. doi: 10.1073/pnas.0507669103. Epub 2006 Mar 6. Proc Natl Acad Sci U S A. 2006. PMID: 16537507 Free PMC article.
-
A conserved major groove antideterminant for Saccharomyces cerevisiae RNase III recognition.Biochemistry. 2005 Mar 22;44(11):4181-7. doi: 10.1021/bi047483u. Biochemistry. 2005. PMID: 15766245
-
Life without RNAi: noncoding RNAs and their functions in Saccharomyces cerevisiae.Biochem Cell Biol. 2009 Oct;87(5):767-79. doi: 10.1139/O09-043. Biochem Cell Biol. 2009. PMID: 19898526 Review.
-
SnoRNP biogenesis meets Pre-mRNA splicing.Mol Cell. 2006 Sep 15;23(6):775-6. doi: 10.1016/j.molcel.2006.08.023. Mol Cell. 2006. PMID: 16973429 Review.
Cited by
-
tRNA 3' processing in yeast involves tRNase Z, Rex1, and Rrp6.RNA. 2014 Jan;20(1):115-30. doi: 10.1261/rna.041467.113. Epub 2013 Nov 18. RNA. 2014. PMID: 24249226 Free PMC article.
-
Loss of the Yeast SR Protein Npl3 Alters Gene Expression Due to Transcription Readthrough.PLoS Genet. 2015 Dec 22;11(12):e1005735. doi: 10.1371/journal.pgen.1005735. eCollection 2015 Dec. PLoS Genet. 2015. PMID: 26694144 Free PMC article.
-
Retrograde nuclear import of tRNA precursors is required for modified base biogenesis in yeast.Proc Natl Acad Sci U S A. 2011 Jun 28;108(26):10502-7. doi: 10.1073/pnas.1105645108. Epub 2011 Jun 13. Proc Natl Acad Sci U S A. 2011. PMID: 21670254 Free PMC article.
-
Analysis of Escherichia coli RNase E and RNase III activity in vivo using tiling microarrays.Nucleic Acids Res. 2011 Apr;39(8):3188-203. doi: 10.1093/nar/gkq1242. Epub 2010 Dec 11. Nucleic Acids Res. 2011. PMID: 21149258 Free PMC article.
-
A homolog of lariat-debranching enzyme modulates turnover of branched RNA.RNA. 2014 Aug;20(8):1337-48. doi: 10.1261/rna.044602.114. Epub 2014 Jun 11. RNA. 2014. PMID: 24919400 Free PMC article.
References
-
- Sherman F. Getting started with yeast. Methods Enzymol. 2002;350:3–41. - PubMed
-
- Venema J., Tollervey D. Ribosome synthesis in Saccharomyces cerevisiae. Annu. Rev. Genet. 1999;33:261–311. - PubMed
-
- Decatur W.A., Fournier M.J. rRNA modifications and ribosome function. Trends Biochem. Sci. 2002;27:344–351. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
