

Opposing roles for DNA structure-specific proteins Rad1, Msh2, Msh3, and Sgs1 in yeast gene targeting
- PMID: 15920474
- PMCID: PMC1150892
- DOI: 10.1038/sj.emboj.7600698
Opposing roles for DNA structure-specific proteins Rad1, Msh2, Msh3, and Sgs1 in yeast gene targeting
Abstract
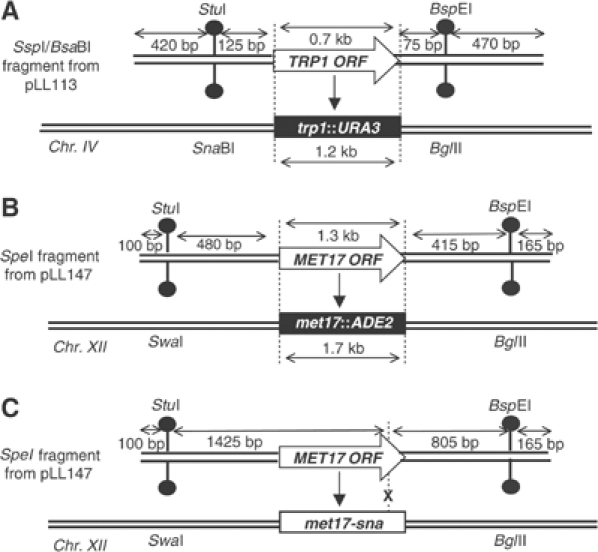
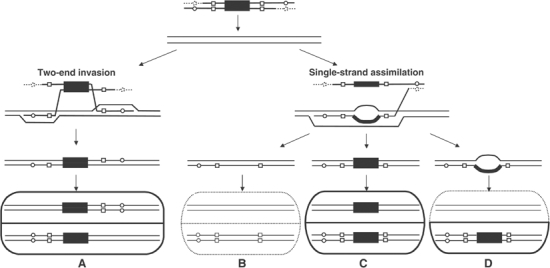
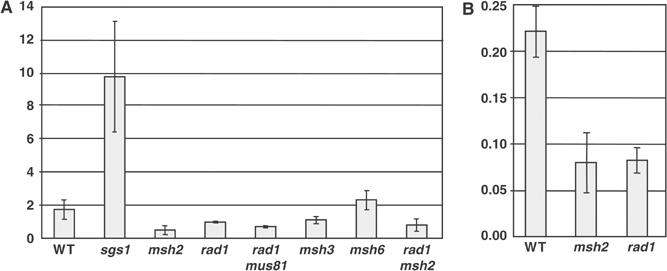
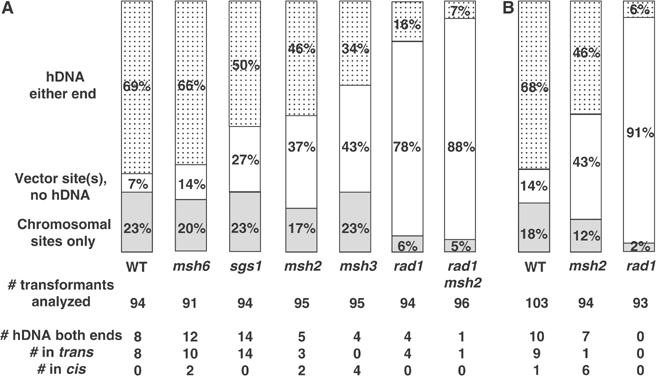
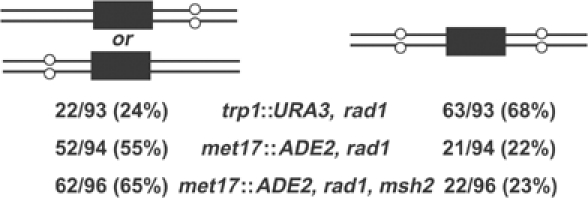
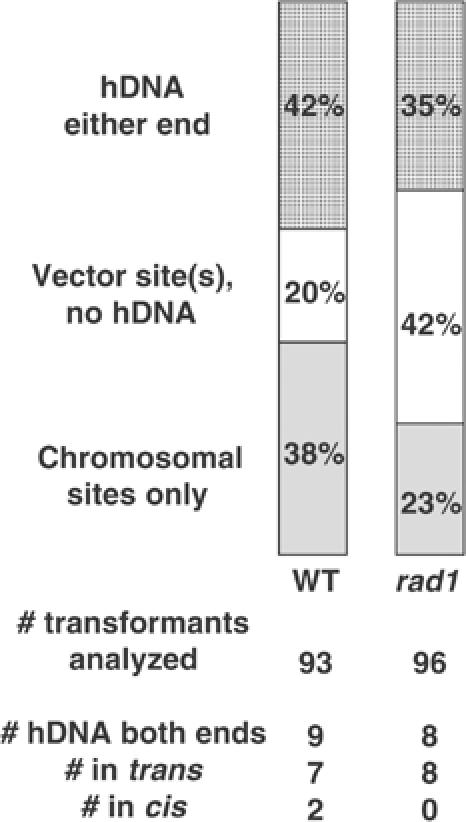
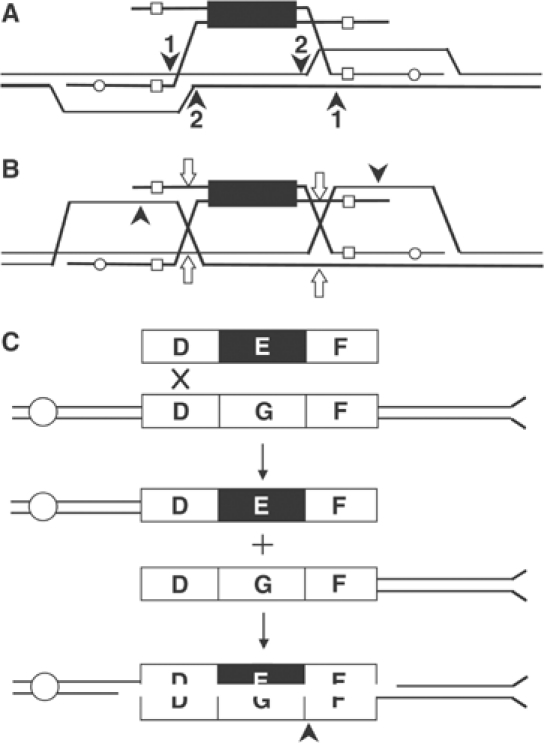
Targeted gene replacement (TGR) in yeast and mammalian cells is initiated by the two free ends of the linear targeting molecule, which invade their respective homologous sequences in the chromosome, leading to replacement of the targeted locus with a selectable gene from the targeting DNA. To study the postinvasion steps in recombination, we examined the effects of DNA structure-specific proteins on TGR frequency and heteroduplex DNA formation. In strains deleted of RAD1, MSH2, or MSH3, we find that the frequency of TGR is reduced and the mechanism of TGR is altered while the reverse is true for deletion of SGS1, suggesting that Rad1 and Msh2:Msh3 facilitate TGR while Sgs1 opposes it. The altered mechanism of TGR in the absence of Msh2:Msh3 and Rad1 reveals a separate role for these proteins in suppressing an alternate gene replacement pathway in which incorporation of both homology regions from a single strand of targeting DNA into heteroduplex with the targeted locus creates a mismatch between the selectable gene on the targeting DNA and the targeted gene in the chromosome.
Figures
References
-
- Bardwell AJ, Bardwell L, Tomkinson AE, Friedberg EC (1994) Specific cleavage of model recombination and repair intermediates by the yeast Rad1–Rad10 DNA endonuclease. Science 265: 2082–2085 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
