

The role of salicylic acid in the induction of cell death in Arabidopsis acd11
- PMID: 15923330
- PMCID: PMC1150418
- DOI: 10.1104/pp.105.059303
The role of salicylic acid in the induction of cell death in Arabidopsis acd11
Abstract
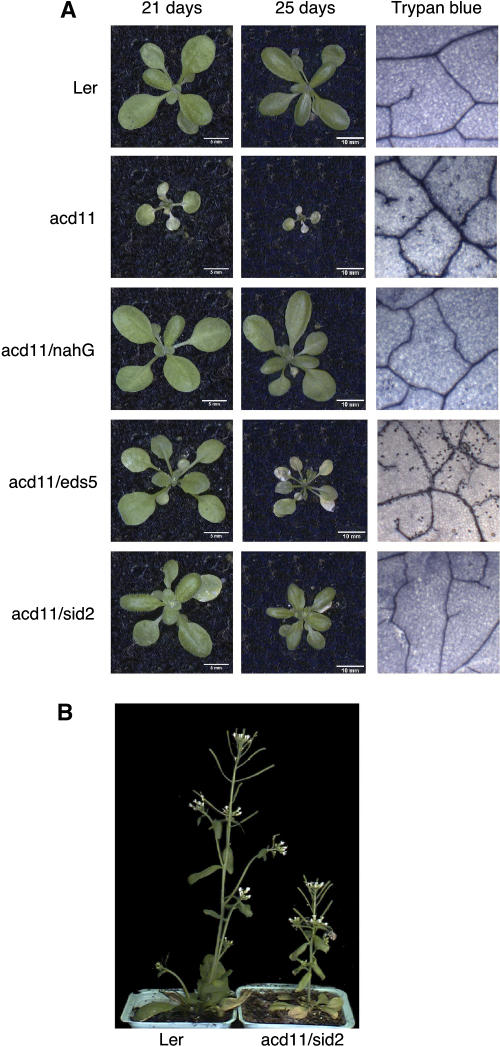
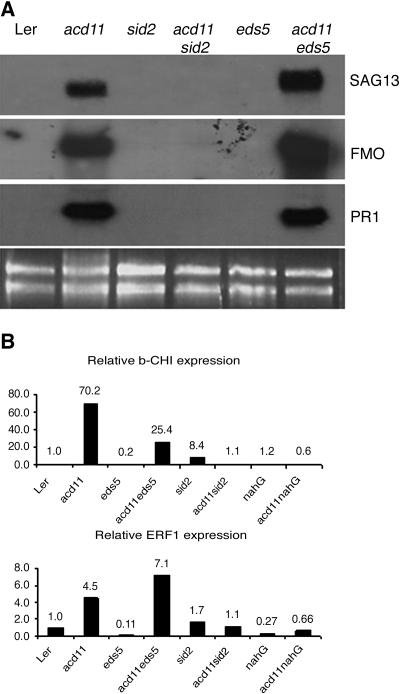
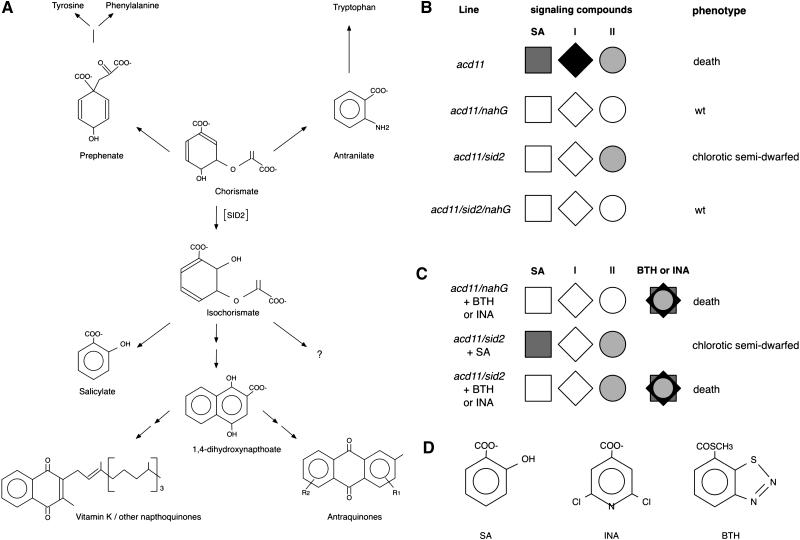
Salicylic acid (SA) is implicated in the induction of programmed cell death (PCD) associated with pathogen defense responses because SA levels increase in response to PCD-inducing infections, and PCD development can be inhibited by expression of salicylate hydroxylase encoded by the bacterial nahG gene. The acd11 mutant of Arabidopsis (Arabidopsis thaliana L. Heynh.) activates PCD and defense responses that are fully suppressed by nahG. To further study the role of SA in PCD induction, we compared phenotypes of acd11/nahG with those of acd11/eds5-1 and acd11/sid2-2 mutants deficient in a putative transporter and isochorismate synthase required for SA biosynthesis. We show that sid2-2 fully suppresses SA accumulation and cell death in acd11, although growth inhibition and premature leaf chlorosis still occur. In addition, application of exogenous SA to acd11/sid2-2 is insufficient to restore cell death. This indicates that isochorismate-derived compounds other than SA are required for induction of PCD in acd11 and that some acd11 phenotypes require NahG-degradable compounds not synthesized via isochorismate.
Figures

Similar articles
-
Knockout of Arabidopsis accelerated-cell-death11 encoding a sphingosine transfer protein causes activation of programmed cell death and defense.Genes Dev. 2002 Feb 15;16(4):490-502. doi: 10.1101/gad.218202. Genes Dev. 2002. PMID: 11850411 Free PMC article.
-
Arabidopsis local resistance to Botrytis cinerea involves salicylic acid and camalexin and requires EDS4 and PAD2, but not SID2, EDS5 or PAD4.Plant J. 2003 Jul;35(2):193-205. doi: 10.1046/j.1365-313x.2003.01794.x. Plant J. 2003. PMID: 12848825
-
Ozone-induced expression of the Arabidopsis FAD7 gene requires salicylic acid, but not NPR1 and SID2.Plant Cell Physiol. 2006 Mar;47(3):355-62. doi: 10.1093/pcp/pci253. Epub 2006 Jan 13. Plant Cell Physiol. 2006. PMID: 16415067
-
Biosynthesis of salicylic acid in plants.Plant Signal Behav. 2009 Jun;4(6):493-6. doi: 10.4161/psb.4.6.8392. Epub 2009 Jun 12. Plant Signal Behav. 2009. PMID: 19816125 Free PMC article. Review.
-
Dead cells do tell tales.Curr Opin Plant Biol. 1998 Dec;1(6):480-5. doi: 10.1016/s1369-5266(98)80039-3. Curr Opin Plant Biol. 1998. PMID: 10066637 Review.
Cited by
-
Lazarus1, a DUF300 protein, contributes to programmed cell death associated with Arabidopsis acd11 and the hypersensitive response.PLoS One. 2010 Sep 7;5(9):e12586. doi: 10.1371/journal.pone.0012586. PLoS One. 2010. PMID: 20830211 Free PMC article.
-
PROHIBITIN3 Forms Complexes with ISOCHORISMATE SYNTHASE1 to Regulate Stress-Induced Salicylic Acid Biosynthesis in Arabidopsis.Plant Physiol. 2018 Mar;176(3):2515-2531. doi: 10.1104/pp.17.00941. Epub 2018 Feb 1. Plant Physiol. 2018. PMID: 29438088 Free PMC article.
-
Autoimmunity in plants.Planta. 2018 Oct;248(4):751-767. doi: 10.1007/s00425-018-2956-0. Epub 2018 Jul 25. Planta. 2018. PMID: 30046903 Review.
-
The E3 Ubiquitin Ligase ATL9 Affects Expression of Defense Related Genes, Cell Death and Callose Deposition in Response to Fungal Infection.Pathogens. 2022 Jan 5;11(1):68. doi: 10.3390/pathogens11010068. Pathogens. 2022. PMID: 35056016 Free PMC article.
-
Human GLTP and mutant forms of ACD11 suppress cell death in the Arabidopsis acd11 mutant.FEBS J. 2008 Sep;275(17):4378-88. doi: 10.1111/j.1742-4658.2008.06584.x. Epub 2008 Jul 24. FEBS J. 2008. PMID: 18657186 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
