

Oct-3/4 maintains the proliferative embryonic stem cell state via specific binding to a variant octamer sequence in the regulatory region of the UTF1 locus
- PMID: 15923625
- PMCID: PMC1140574
- DOI: 10.1128/MCB.25.12.5084-5094.2005
Oct-3/4 maintains the proliferative embryonic stem cell state via specific binding to a variant octamer sequence in the regulatory region of the UTF1 locus
Abstract
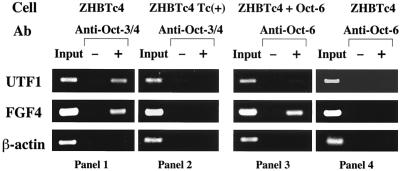
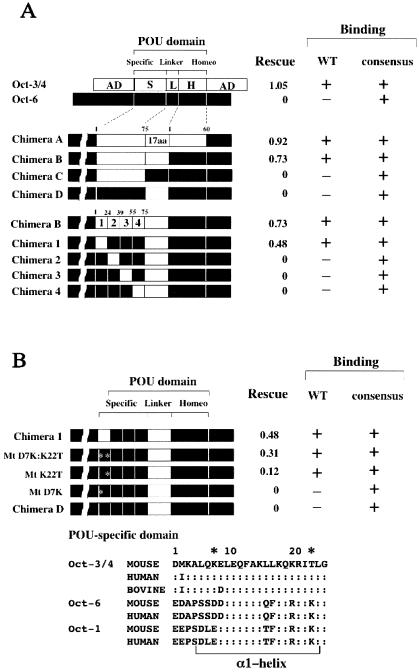
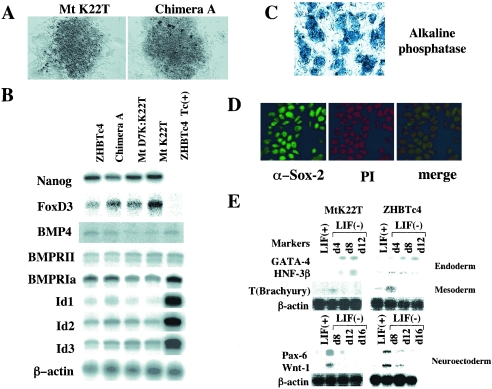
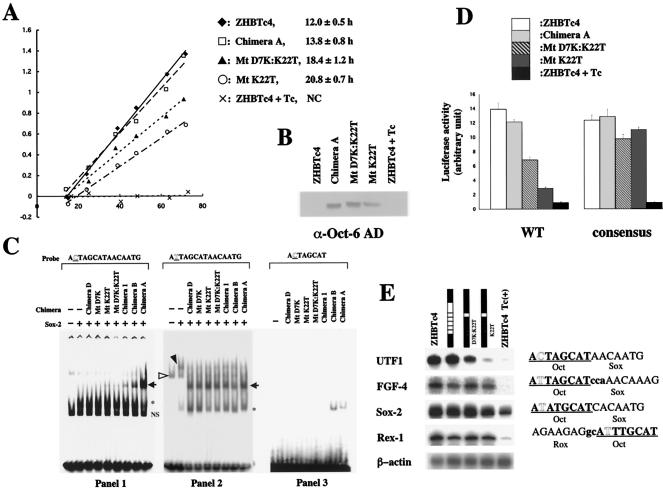
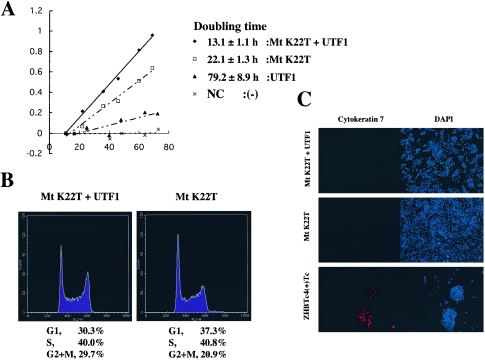
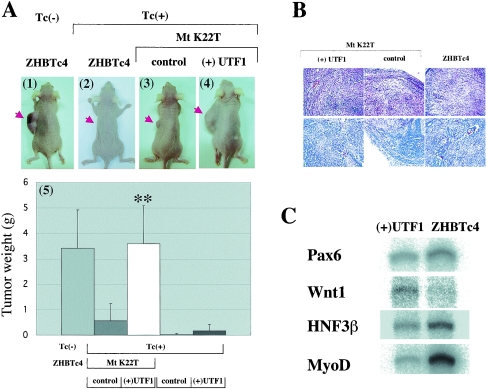
The POU transcription factor Oct-3/4 has been shown to be critical for maintaining embryonic stem (ES) cell character. However, the molecular mechanisms underlying its function remain elusive. We have previously shown that among the POU transcription factor family of proteins, Oct-3/4 alone is able to bind to the regulatory region of the UTF1 gene bearing a variant octamer sequence together with Sox-2. Here, we demonstrate using Oct-3/4-Oct-6 chimeras that there is a precise correlation between the ability of proteins to form a complex on the UTF1 enhancer with Sox-2 and the ability to maintain the stem cell state in ES cells. Different chimeric proteins show differential abilities to form a Sox-2-containing complex on the UTF1 regulatory region, with a decrease in efficiency of the complex formation accompanied by a decrease in the level of UTF1 expression and the rate of cell proliferation. Overexpression of UTF1 in these slow-growing cells was able to restore their proliferation rate to wild-type levels. Moreover, UTF1 was also observed to have an effect on teratoma formation. These results suggest a molecular pathway by which Oct-3/4 induces rapid proliferation and tumorigenic properties of ES cells through activation of the UTF1 gene.
Figures
References
-
- Antonchuk, J., G. Sauvageau, and R. K. Humphries. 2002. HOXB4-induced expansion of adult hematopoietic stem cells ex vivo. Cell 109:39-45. - PubMed
-
- Ben-Shushan, E., J. R. Thompson, L. J. Gudas, and Y. Bergman. 1998. Rex-1, a gene encoding a transcription factor expressed in the early embryo, is regulated via Oct-3/4 and Oct-6 binding to an octamer site and a novel protein, Rox-1, binding to an adjacent site. Mol. Cell. Biol. 18:1866-1878. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous