

Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts
- PMID: 15923655
- PMCID: PMC1283079
- DOI: 10.1242/jcs.02371
Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts
Abstract
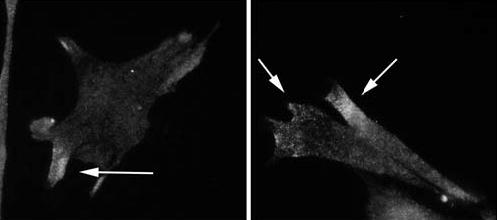
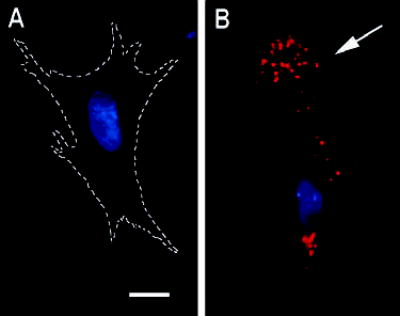
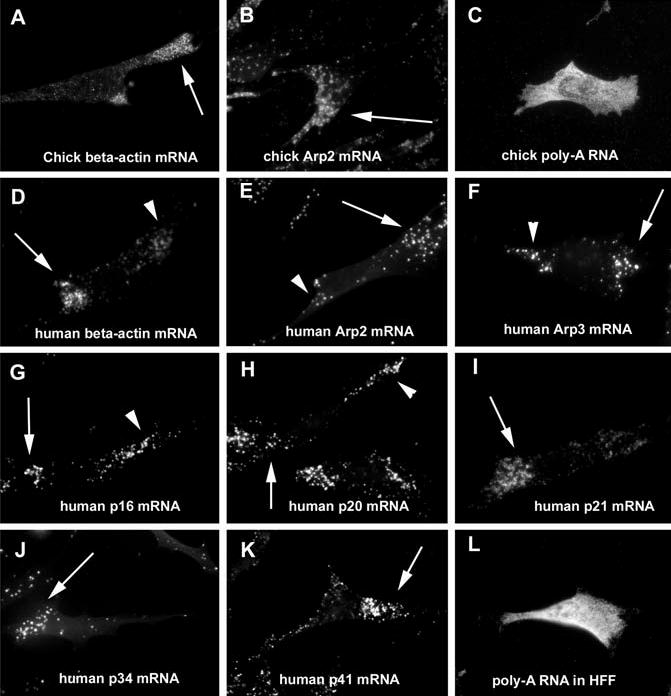
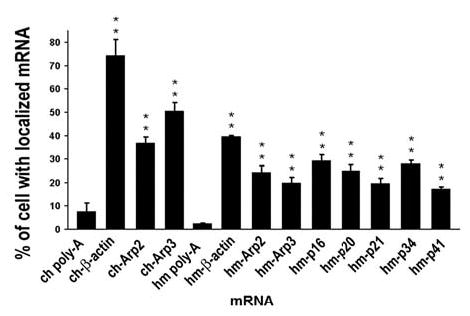
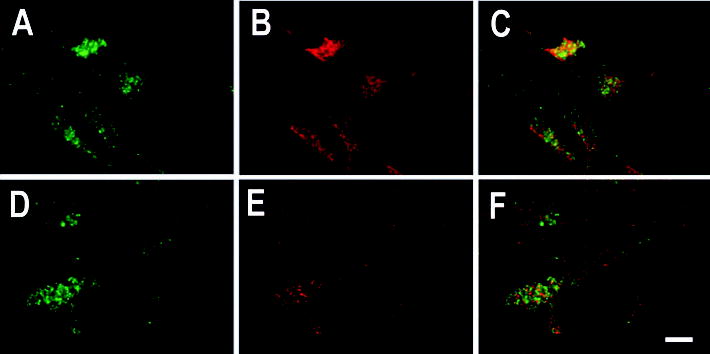
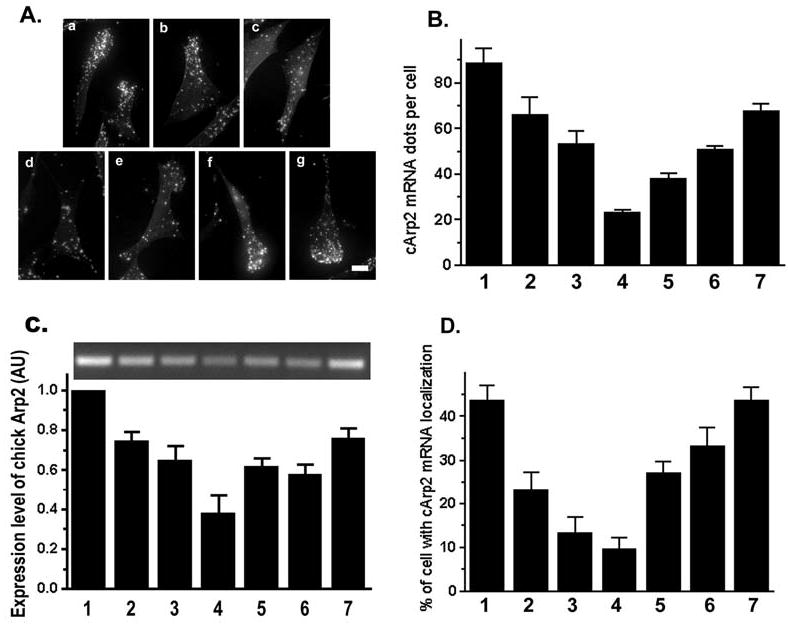
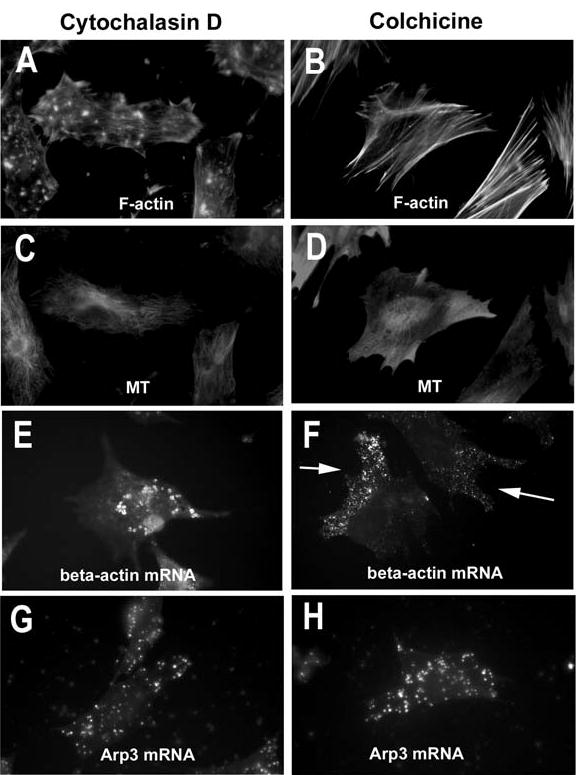
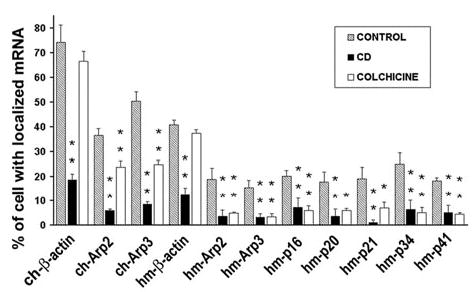
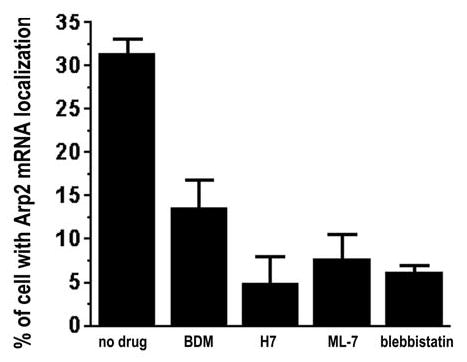
The actin-related protein 2/3 (Arp2/3) complex is a crucial actin polymerization nucleator and is localized to the leading protrusions of migrating cells. However, how the multiprotein complex is targeted to the protrusions remains unknown. Here, we demonstrate that mRNAs for the seven subunits of the Arp2/3 complex are localized to the protrusions in fibroblasts, supporting a hypothesis that the Arp2/3 complex is targeted to its site of function by mRNA localization. Depletion of serum from culture medium inhibits Arp2/3-complex mRNA localization to the protrusion, whereas serum stimulation leads to significant mRNA localization within 30 minutes. The effect of serum suggests that Arp2/3-complex mRNA localization is a cellular response to extracellular stimuli. The localization of the Arp2/3 complex mRNAs is dependent on both actin filaments and microtubules, because disruption of either cytoskeletal system (with cytochalasin D and colchicine, respectively) inhibited the localization of all seven subunit mRNAs. In addition, myosin inhibitors significantly inhibit Arp2 mRNA localization in chicken embryo fibroblasts, suggesting a myosin motor dependent mechanism for Arp2/3-complex mRNA localization.
Figures
Comment in
-
Localizing cellular housekeeping.Nat Rev Mol Cell Biol. 2011 Dec;12(12):771. doi: 10.1038/nrm3206. Nat Rev Mol Cell Biol. 2011. PMID: 21971042 No abstract available.
References
-
- Bailly M, Ichetovkin I, Grant W, Zebda N, Machesky LM, Segall JE, Condeelis J. The F-actin side binding activity of the Arp2/3 complex is essential for actin nucleation and lamellipod extension. Curr Biol. 2001;11:620–625. - PubMed
-
- Barbarese E, Koppel DE, Deutscher MP, Smith CL, Ainger K, Morgan F, Carson JH. Protein translation components are colocalized in granules in oligodendrocytes. J Cell Sci. 1995;108:2781–2790. - PubMed
-
- Bassell GJ, Oleynikov Y, Singer RH. The travels of mRNAs through all cells large and small. FASEB J. 1999;13:447–454. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
