

Recruitment of Gr-1+ monocytes is essential for control of acute toxoplasmosis
- PMID: 15928200
- PMCID: PMC2213275
- DOI: 10.1084/jem.20050054
Recruitment of Gr-1+ monocytes is essential for control of acute toxoplasmosis
Abstract
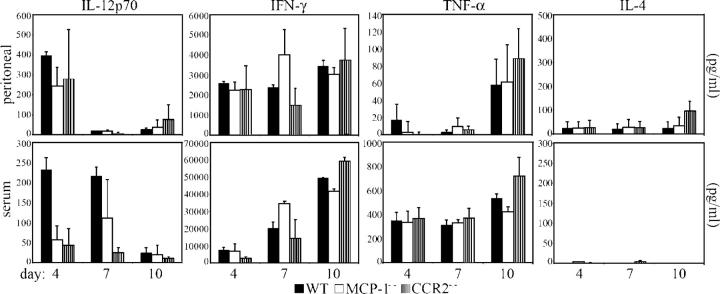
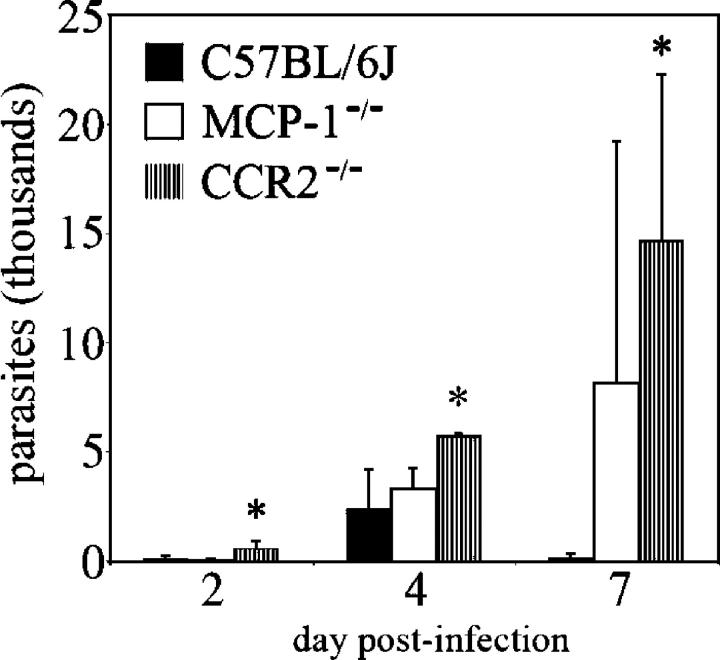
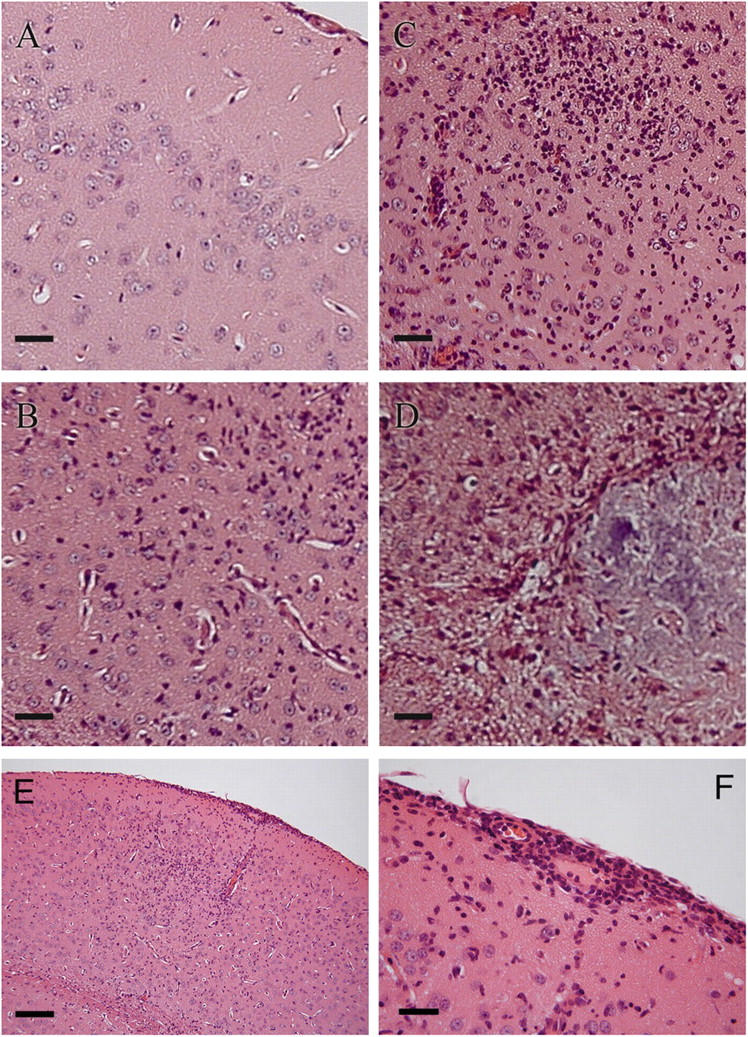
Circulating murine monocytes comprise two largely exclusive subpopulations that are responsible for seeding normal tissues (Gr-1-/CCR2-/CX3CR1high) or responding to sites of inflammation (Gr-1+/CCR2+/CX3CR1lo). Gr-1+ monocytes are recruited to the site of infection during the early stages of immune response to the intracellular pathogen Toxoplasma gondii. A murine model of toxoplasmosis was thus used to examine the importance of Gr-1+ monocytes in the control of disseminated parasitic infection in vivo. The recruitment of Gr-1+ monocytes was intimately associated with the ability to suppress early parasite replication at the site of inoculation. Infection of CCR2-/- and MCP-1-/- mice with typically nonlethal, low doses of T. gondii resulted in the abrogated recruitment of Gr-1+ monocytes. The failure to recruit Gr-1+ monocytes resulted in greatly enhanced mortality despite the induction of normal Th1 cell responses leading to high levels of IL-12, TNF-alpha, and IFN-gamma. The profound susceptibility of CCR2-/- mice establishes Gr-1+ monocytes as necessary effector cells in the resistance to acute toxoplasmosis and suggests that the CCR2-dependent recruitment of Gr-1+ monocytes may be an important general mechanism for resistance to intracellular pathogens.
Figures
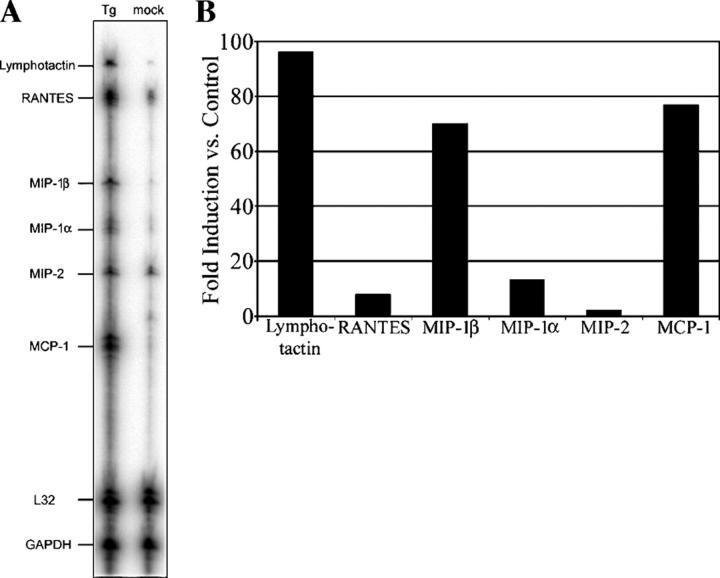
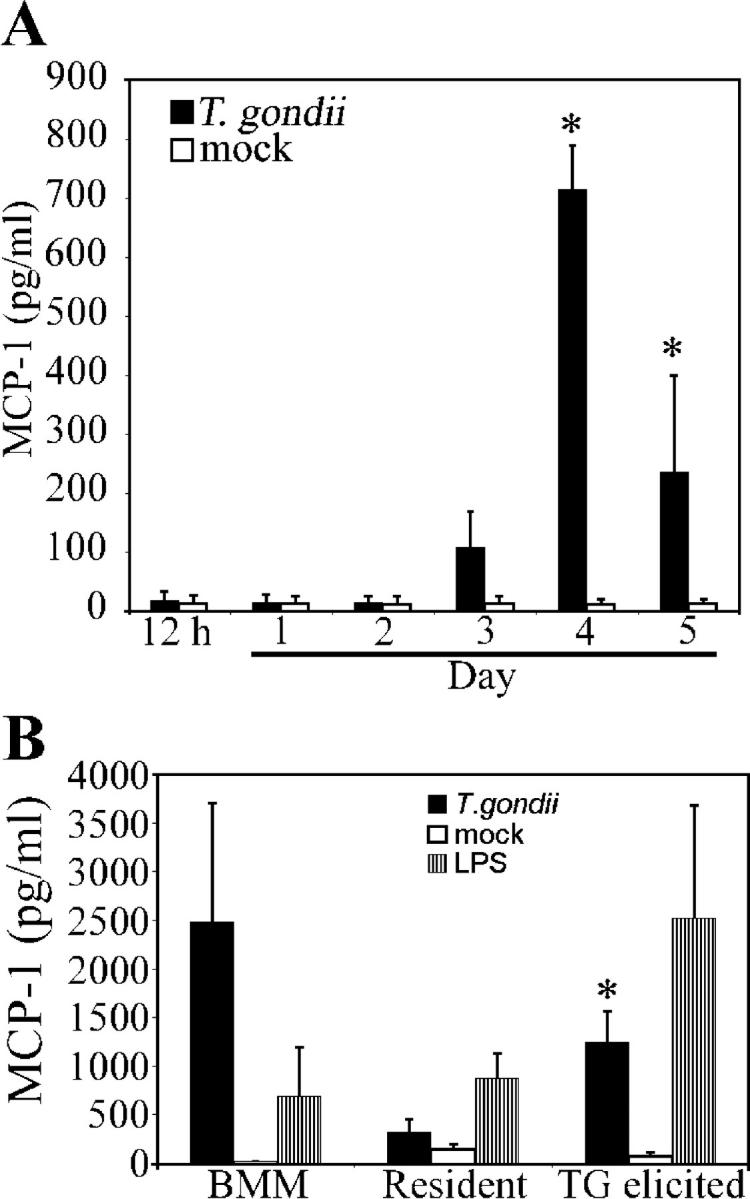
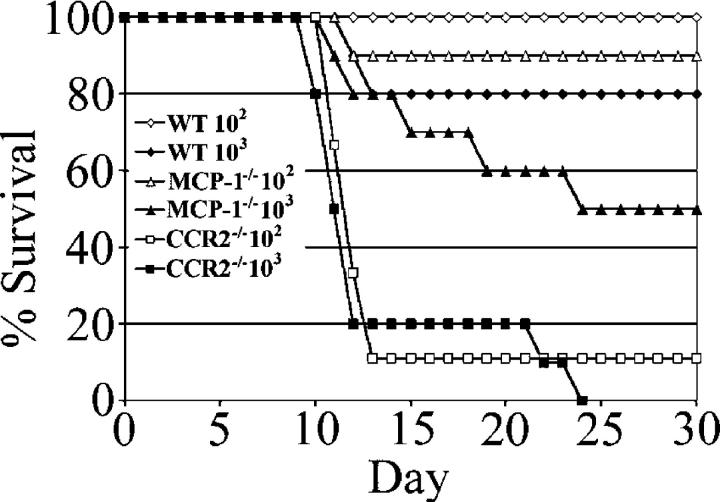
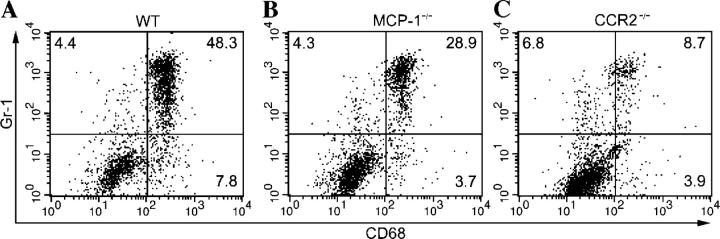
References
-
- Muller, W.A., and G.J. Randolph. 1999. Migration of leukocytes across endothelium and beyond: molecules involved in the transmigration and fate of monocytes. J. Leukoc. Biol. 66:698–704. - PubMed
-
- Passlick, B., D. Flieger, and H.W. Ziegler-Heitbrock. 1989. Identification and characterization of a novel monocytes subpopulation in human peripheral blood. Blood. 74:2527–2534. - PubMed
-
- Geissmann, F., S. Jung, and D.R. Littman. 2003. Blood monocytes consist of two principle subsets with distinct migratory properties. Immunity. 19:71–82. - PubMed
-
- Henderson, R.B., J.A.R. Hobbs, M. Mathies, and N. Hogg. 2003. Rapid recruitment of inflammatory monocytes is independent of neutrophil migration. Blood. 102:328–335. - PubMed
-
- Taylor, P.R., and S. Gordon. 2003. Pattern recognition receptors and differentiation antigens define murine myeloid cell heterogeneity ex vivo. Eur. J. Immunol. 33:2090–2097. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
