

Inp1p is a peroxisomal membrane protein required for peroxisome inheritance in Saccharomyces cerevisiae
- PMID: 15928207
- PMCID: PMC2171609
- DOI: 10.1083/jcb.200503083
Inp1p is a peroxisomal membrane protein required for peroxisome inheritance in Saccharomyces cerevisiae
Abstract
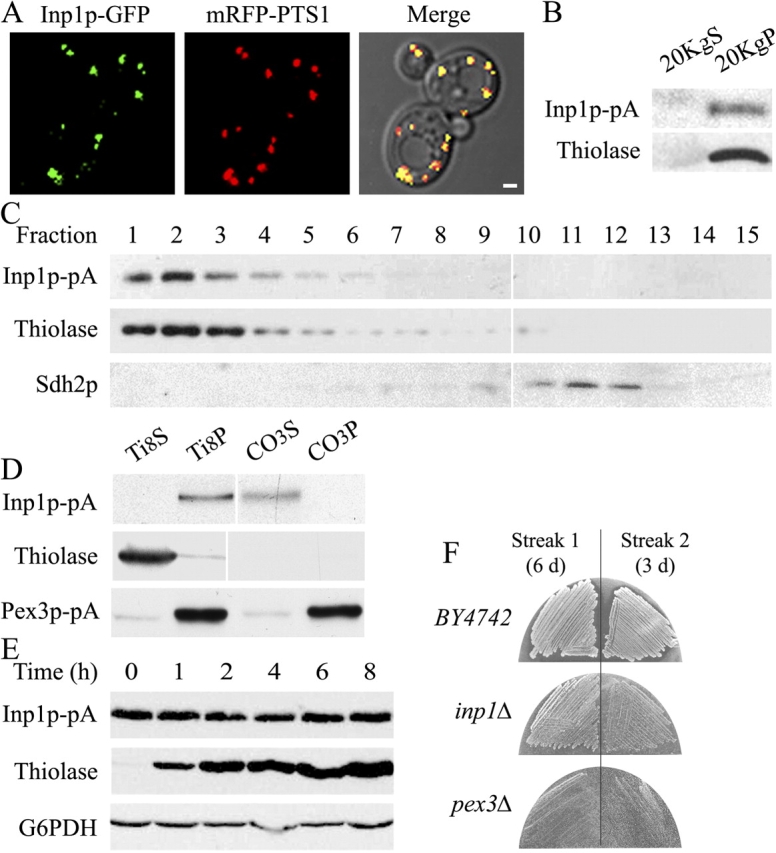
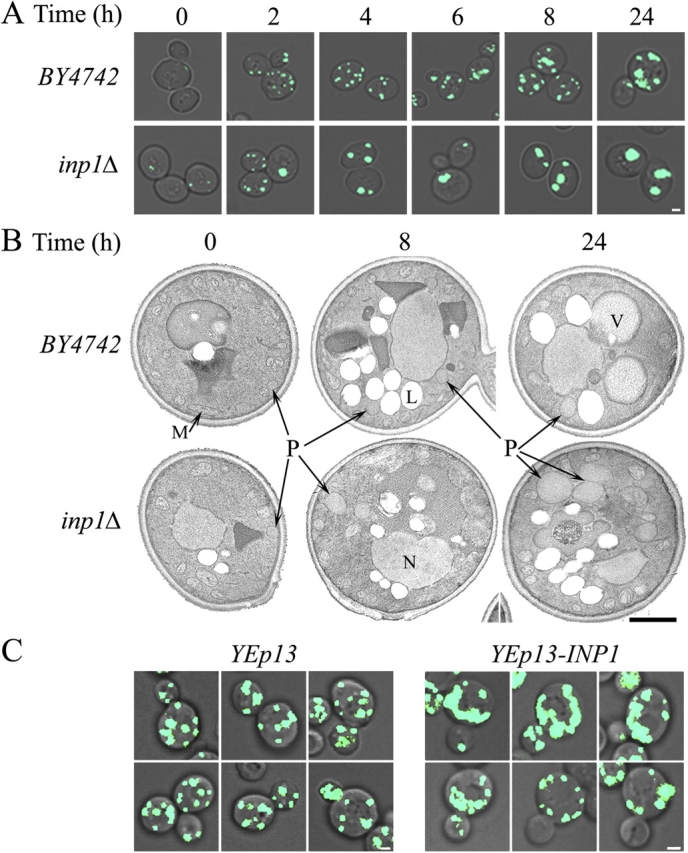
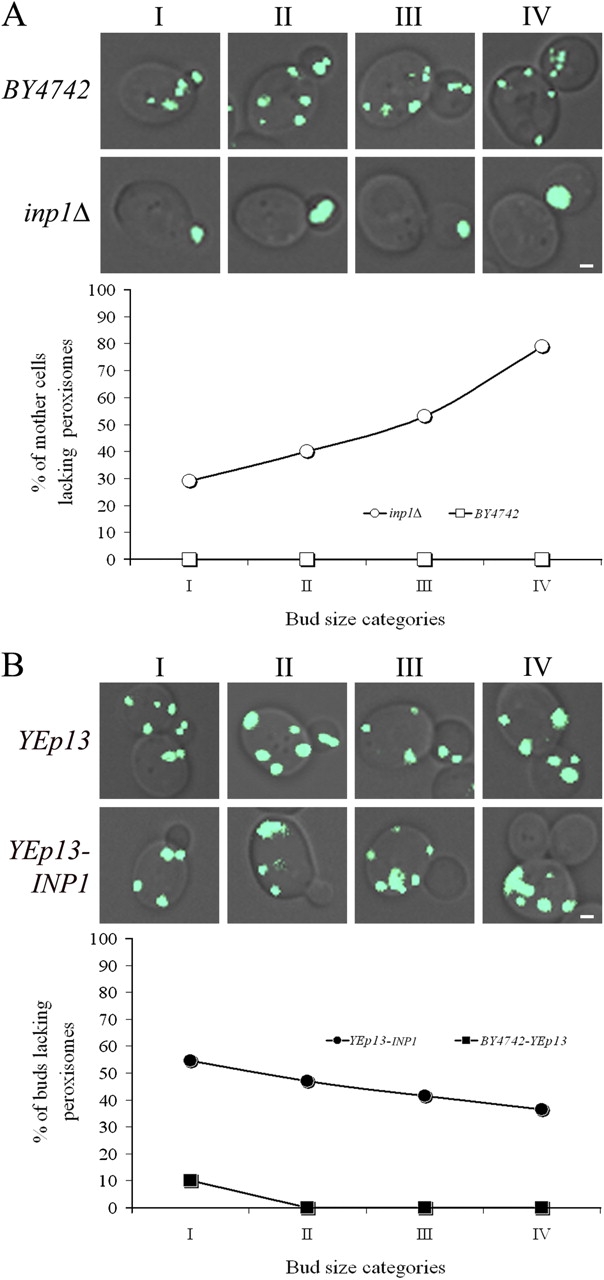
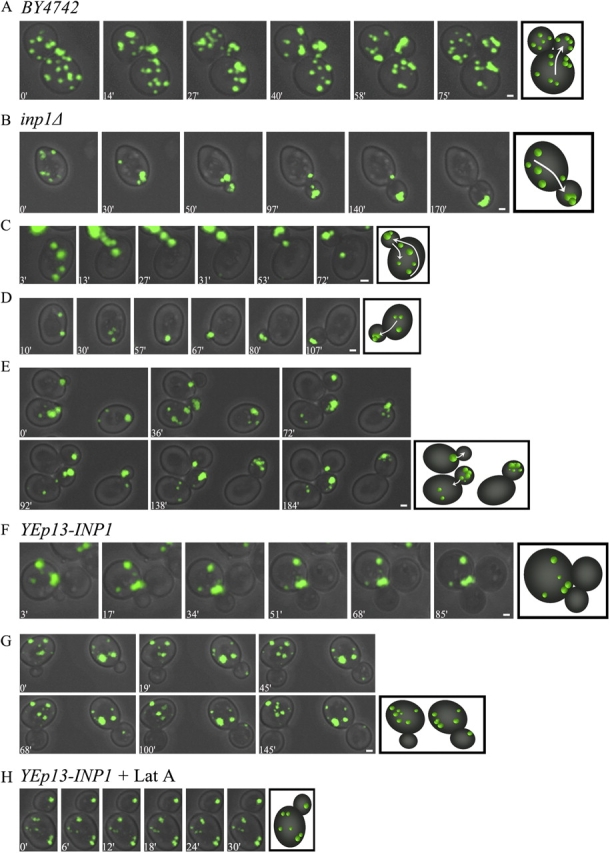
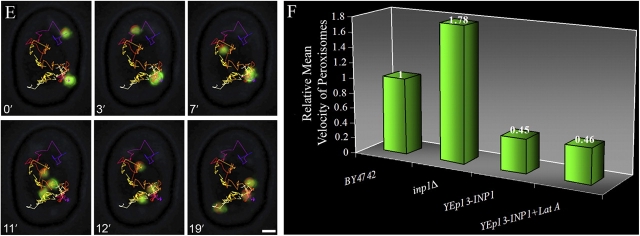
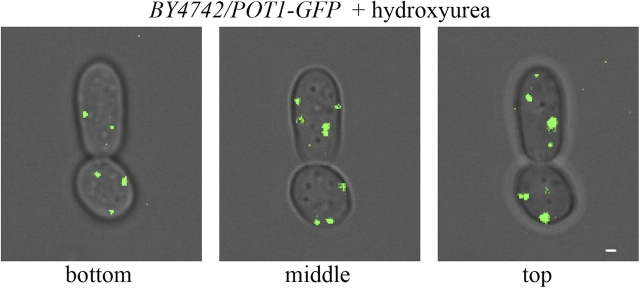
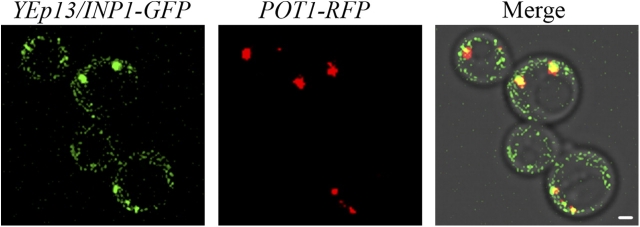
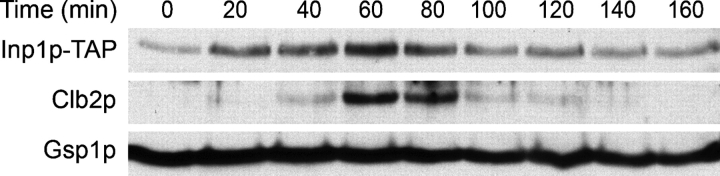
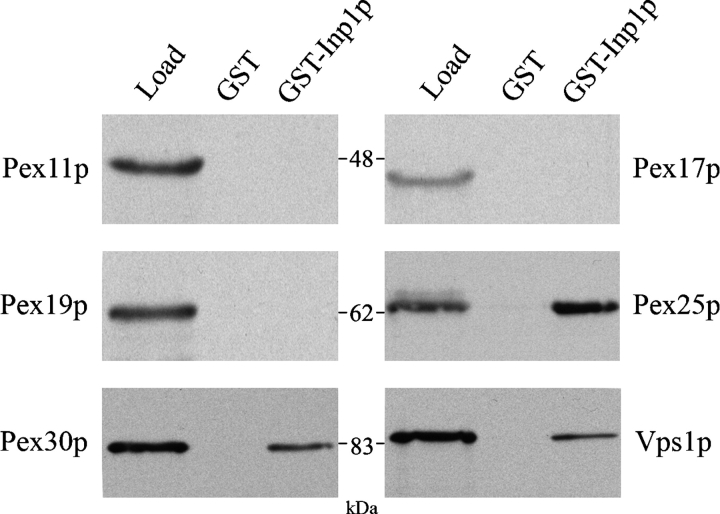
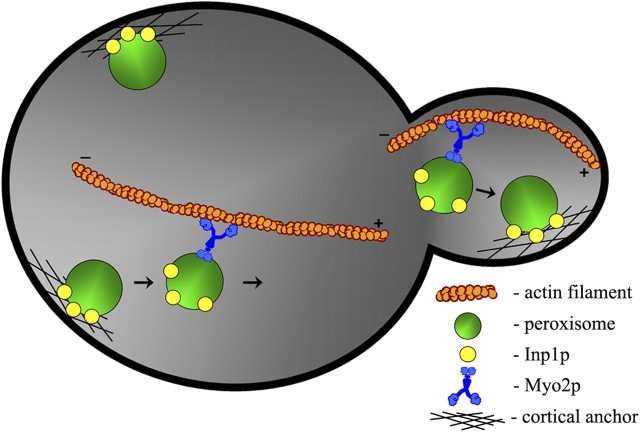
Cells have evolved molecular mechanisms for the efficient transmission of organelles during cell division. Little is known about how peroxisomes are inherited. Inp1p is a peripheral membrane protein of peroxisomes of Saccharomyces cerevisiae that affects both the morphology of peroxisomes and their partitioning during cell division. In vivo 4-dimensional video microscopy showed an inability of mother cells to retain a subset of peroxisomes in dividing cells lacking the INP1 gene, whereas cells overexpressing INP1 exhibited immobilized peroxisomes that failed to be partitioned to the bud. Overproduced Inp1p localized to both peroxisomes and the cell cortex, supporting an interaction of Inp1p with specific structures lining the cell periphery. The levels of Inp1p vary with the cell cycle. Inp1p binds Pex25p, Pex30p, and Vps1p, which have been implicated in controlling peroxisome division. Our findings are consistent with Inp1p acting as a factor that retains peroxisomes in cells and controls peroxisome division. Inp1p is the first peroxisomal protein directly implicated in peroxisome inheritance.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
