

Peroxin 5-peroxin 14 association in the protozoan Leishmania donovani involves a novel protein-protein interaction motif
- PMID: 15929724
- PMCID: PMC1237144
- DOI: 10.1042/BJ20050328
Peroxin 5-peroxin 14 association in the protozoan Leishmania donovani involves a novel protein-protein interaction motif
Abstract
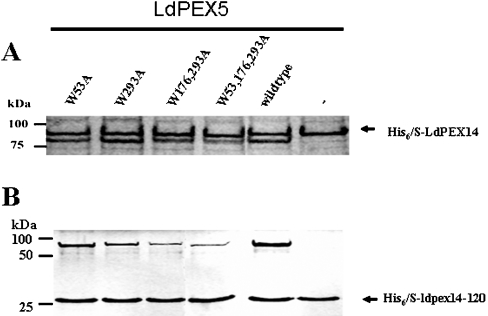
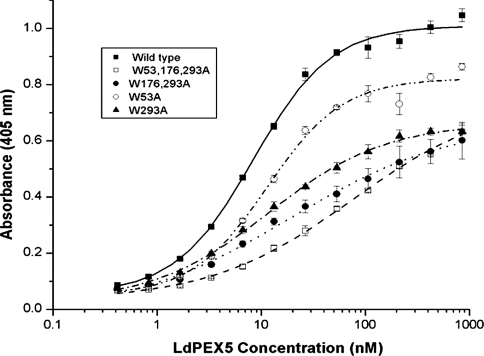
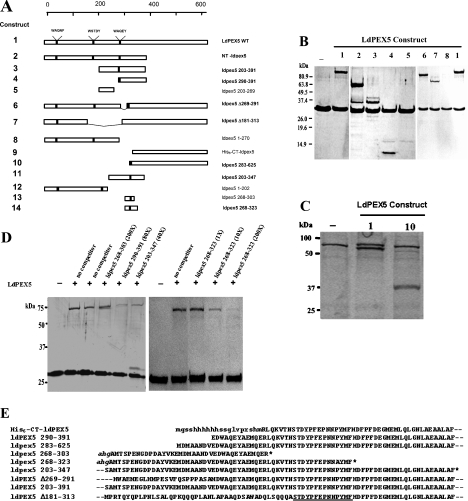
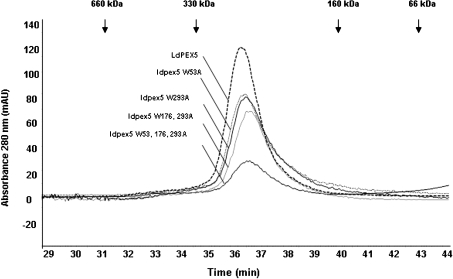
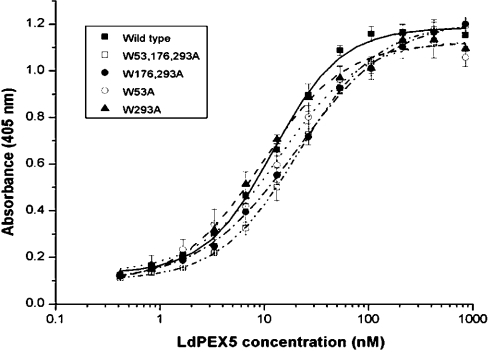
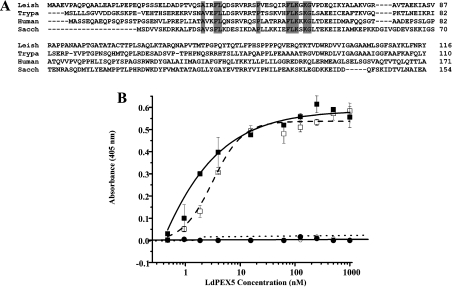
Import of proteins with a PTS1 (peroxisomal targeting signal 1) into the Leishmania glycosomal organelle involves docking of a PTS1-laden LdPEX5 [Leishmania donovani PEX5 (peroxin 5)] receptor to LdPEX14 on the surface of the glycosomal membrane. In higher eukaryotes, the PEX5-PEX14 interaction is mediated by a conserved diaromatic WXXXY/F motif. Site-directed and deletion mutageneses of the three WXXXY/F repeats in LdPEX5 did not abolish the LdPEX5-LdPEX14 association. Analysis of the equilibrium dissociation constant (K(d)) revealed that ldpex5-W53A (Trp53-->Ala), ldpex5-W293A, ldpex5-W176,293A and ldpex5-W53,176,293A mutant receptors were capable of binding LdPEX14 with affinities comparable with wild-type LdPEX5. That the diaromatic motifs were not required for the LdPEX5-LdPEX14 interaction was further verified by deletion analysis that showed that ldpex5 deletion mutants or ldpex5 fragments lacking the WXXXY/F motifs retained LdPEX14 binding activity. Mapping studies of LdPEX5 indicated that the necessary elements required for LdPEX14 association were localized to a region between residues 290 and 323. Finally, mutational analysis of LdPEX14 confirmed that residues 23-63, which encompass the conserved signature sequence AX2FLX7SPX6FLKGKGL/V present in all PEX14 proteins, are essential for LdPEX5 binding.
Figures

References
-
- Dacks J. B., Doolittle W. F. Reconstructing/deconstructing the earliest eukaryotes: how comparative genomics can help. Cell (Cambridge, Mass.) 2001;107:419–425. - PubMed
-
- Zeiner G. M., Sturm N. R., Campbell D. A. The Leishmania tarentolae spliced leader contains determinants for association with polysomes. J. Biol. Chem. 2003;278:38269–38275. - PubMed
-
- Kable M. L., Heidmann S., Stuart K. D. RNA editing: getting U into RNA. Trends Biochem. Sci. 1997;22:162–168. - PubMed
-
- Englund P. T., Hajduk S. L., Marini J. C. The molecular biology of trypanosomes. Annu. Rev. Biochem. 1982;51:695–726. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials