

A monoclonal antibody that inhibits mycobacterial DNA gyrase by a novel mechanism
- PMID: 15930158
- PMCID: PMC1142348
- DOI: 10.1093/nar/gki622
A monoclonal antibody that inhibits mycobacterial DNA gyrase by a novel mechanism
Abstract
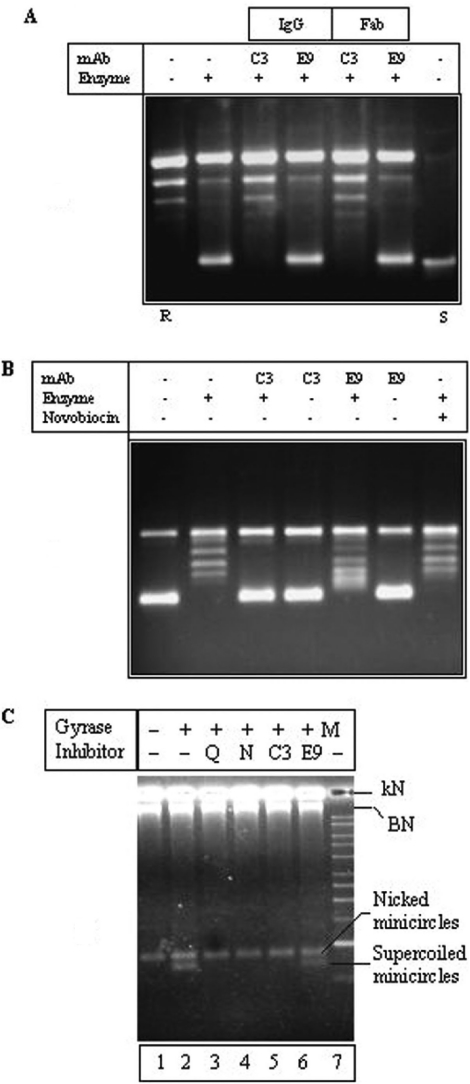
DNA gyrase is a DNA topoisomerase indispensable for cellular functions in bacteria. We describe a novel, hitherto unknown, mechanism of specific inhibition of Mycobacterium smegmatis and Mycobacterium tuberculosis DNA gyrase by a monoclonal antibody (mAb). Binding of the mAb did not affect either GyrA-GyrB or gyrase-DNA interactions. More importantly, the ternary complex of gyrase-DNA-mAb retained the ATPase activity of the enzyme and was competent to catalyse DNA cleavage-religation reactions, implying a new mode of action different from other classes of gyrase inhibitors. DNA gyrase purified from fluoroquinolone-resistant strains of M.tuberculosis and M.smegmatis were inhibited by the mAb. The absence of cross-resistance of the drug-resistant enzymes from two different sources to the antibody-mediated inhibition corroborates the new mechanism of inhibition. We suggest that binding of the mAb in the proximity of the primary dimer interface region of GyrA in the heterotetrameric enzyme appears to block the release of the transported segment after strand passage, leading to enzyme inhibition. The specific inhibition of mycobacterial DNA gyrase with the mAb opens up new avenues for designing novel lead molecules for drug discovery and for probing gyrase mechanism.
Figures







Similar articles
-
Fluoroquinolone-gyrase-DNA complexes: two modes of drug binding.J Biol Chem. 2014 May 2;289(18):12300-12. doi: 10.1074/jbc.M113.529164. Epub 2014 Feb 4. J Biol Chem. 2014. PMID: 24497635 Free PMC article.
-
Effect of different classes of inhibitors on DNA gyrase from Mycobacterium smegmatis.J Antimicrob Chemother. 2001 Oct;48(4):479-85. doi: 10.1093/jac/48.4.479. J Antimicrob Chemother. 2001. PMID: 11581225
-
Mycobacterium tuberculosis DNA gyrase: interaction with quinolones and correlation with antimycobacterial drug activity.Antimicrob Agents Chemother. 2004 Apr;48(4):1281-8. doi: 10.1128/AAC.48.4.1281-1288.2004. Antimicrob Agents Chemother. 2004. PMID: 15047530 Free PMC article.
-
Mycobacterium tuberculosis DNA gyrase as a target for drug discovery.Infect Disord Drug Targets. 2007 Jun;7(2):159-68. doi: 10.2174/187152607781001763. Infect Disord Drug Targets. 2007. PMID: 17970226 Review.
-
Gyrase and Topoisomerase IV: Recycling Old Targets for New Antibacterials to Combat Fluoroquinolone Resistance.ACS Infect Dis. 2024 Apr 12;10(4):1097-1115. doi: 10.1021/acsinfecdis.4c00128. Epub 2024 Apr 2. ACS Infect Dis. 2024. PMID: 38564341 Free PMC article. Review.
Cited by
-
The naphthoquinone diospyrin is an inhibitor of DNA gyrase with a novel mechanism of action.J Biol Chem. 2013 Feb 15;288(7):5149-56. doi: 10.1074/jbc.M112.419069. Epub 2012 Dec 28. J Biol Chem. 2013. PMID: 23275348 Free PMC article.
-
Glutamate racemase from Mycobacterium tuberculosis inhibits DNA gyrase by affecting its DNA-binding.Nucleic Acids Res. 2006;34(19):5567-76. doi: 10.1093/nar/gkl704. Epub 2006 Oct 4. Nucleic Acids Res. 2006. PMID: 17020913 Free PMC article.
-
Peptide Inhibition of Topoisomerase IB from Plasmodium falciparum.Mol Biol Int. 2011;2011:854626. doi: 10.4061/2011/854626. Epub 2011 May 4. Mol Biol Int. 2011. PMID: 22091414 Free PMC article.
-
Recent Development of DNA Gyrase Inhibitors: An Update.Mini Rev Med Chem. 2024;24(10):1001-1030. doi: 10.2174/0113895575264264230921080718. Mini Rev Med Chem. 2024. PMID: 37909434 Review.
References
-
- Champoux J.J. DNA topoisomerases: structure, function, and mechanism. Annu. Rev. Biochem. 2001;70:369–413. - PubMed
-
- Heddle J.G., Mitelheiser S., Maxwell A., Thomson N.H. Nucleotide binding to DNA gyrase causes loss of DNA wrap. J. Mol. Biol. 2004;337:597–610. - PubMed
-
- Maxwell A. DNA gyrase as a drug target. Trends Microbiol. 1997;5:102–109. - PubMed
-
- Drlica K., Malik M. Fluoroquinolones: action and resistance. Curr. Top. Med. Chem. 2003;3:249–282. - PubMed
