

Operon formation is driven by co-regulation and not by horizontal gene transfer
- PMID: 15930492
- PMCID: PMC1142471
- DOI: 10.1101/gr.3368805
Operon formation is driven by co-regulation and not by horizontal gene transfer
Abstract
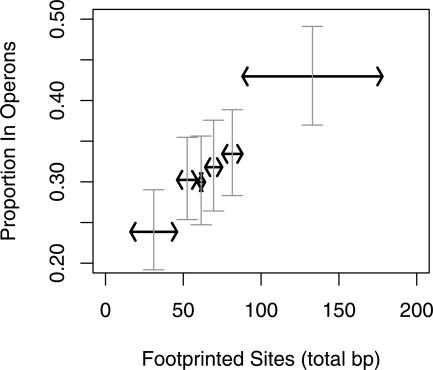
The organization of bacterial genes into operons was originally ascribed to the benefits of co-regulation. More recently, the "selfish operon" model, in which operons are formed by repeated gain and loss of genes, was proposed. Indeed, operons are often subject to horizontal gene transfer (HGT). On the other hand, non-HGT genes are particularly likely to be in operons. To clarify whether HGT is involved in operon formation, we identified recently formed operons in Escherichia coli K12. We show that genes that have homologs in distantly related bacteria but not in close relatives of E. coli--indicating HGT--form new operons at about the same rates as native genes. Furthermore, genes in new operons are no more likely than other genes to have phylogenetic trees that are inconsistent with the species tree. In contrast, essential genes and ubiquitous genes without paralogs--genes believed to undergo HGT rarely--often form new operons. We conclude that HGT is not a cause of operon formation but instead promotes the prevalence of pre-existing operons. To explain operon formation, we propose that new operons reduce the amount of regulatory information required to specify optimal expression patterns and infer that operons should be more likely to evolve than independent promoters when regulation is complex. Consistent with this hypothesis, operons have greater amounts of conserved regulatory sequences than do individually transcribed genes.
Figures


References
-
- Butland, G., Peregrin-Alvarez, J.M., Li, J., Yang, W., Yang, X., Canadien, V., Starostine, A., Richards, D., Beattie, B., Krogan, N., et al. 2005. Interaction network containing conserved and essential protein complexes in Escherichia coli. Nature 433: 531-537. - PubMed
-
- Cherry, J.L. 2003. Genome size and operon content. J. Theor. Biol. 221: 401-410. - PubMed
-
- Dandekar, T., Snel, B., Huynen, M., and Bork, P. 1998. Conservation of gene order: A fingerprint of proteins that physically interact. Trends Biochem. Sci. 23: 324-328. - PubMed
WEB SITE REFERENCES
-
- http://www.r-project.org/; the R statistics package.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases