

Activating transcription factor 3, a stress sensor, activates p53 by blocking its ubiquitination
- PMID: 15933712
- PMCID: PMC1173153
- DOI: 10.1038/sj.emboj.7600712
Activating transcription factor 3, a stress sensor, activates p53 by blocking its ubiquitination
Abstract
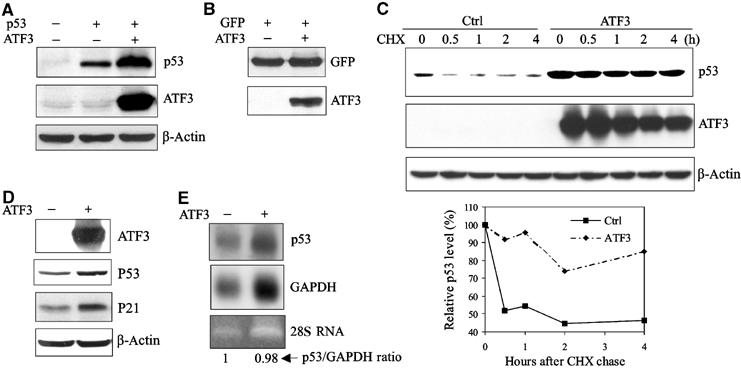
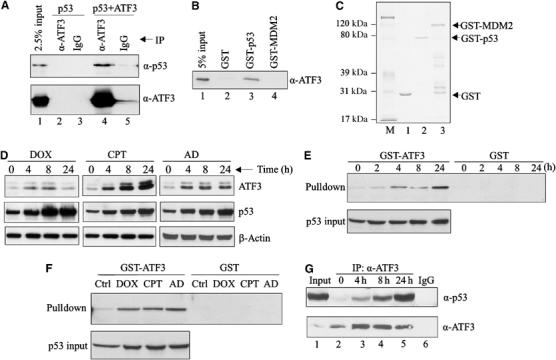
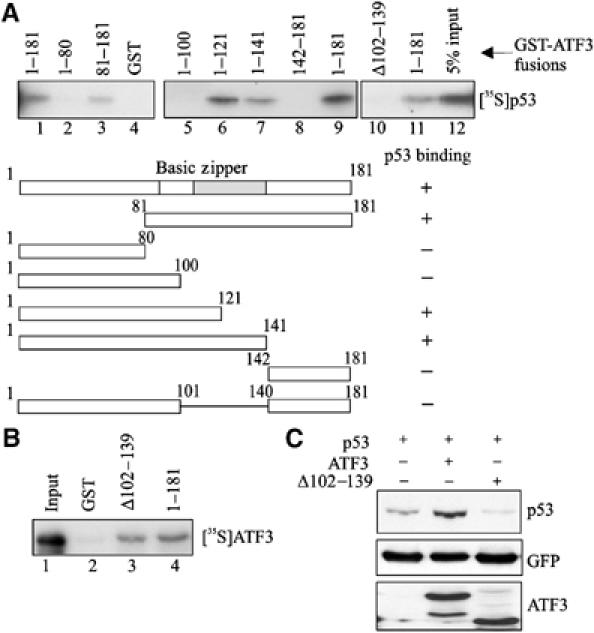
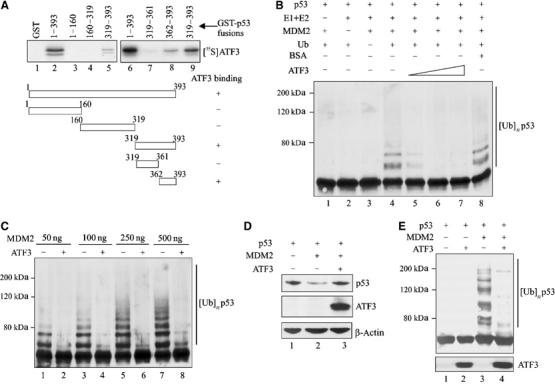
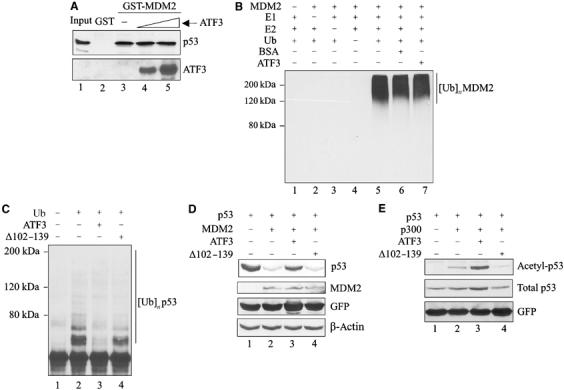
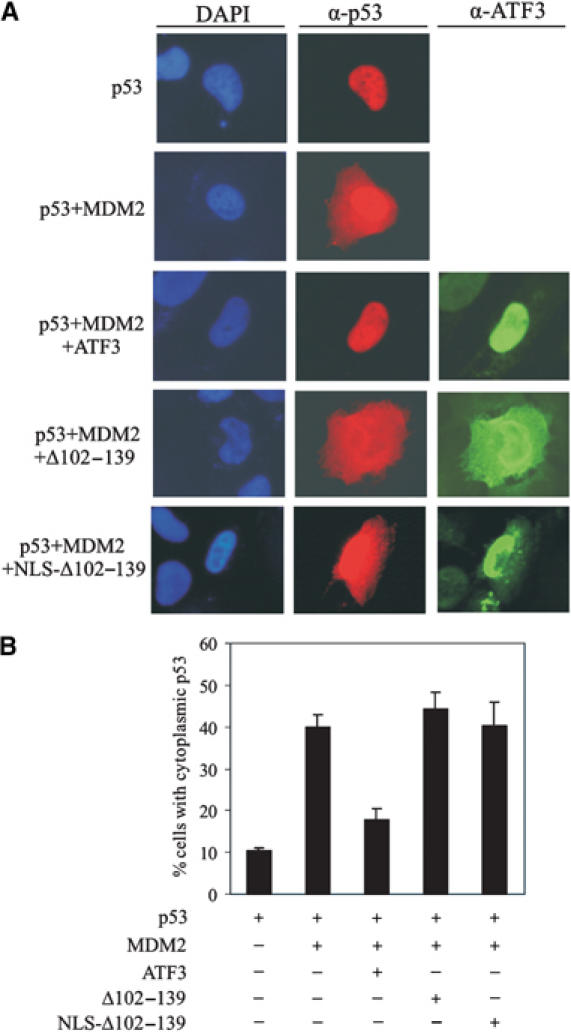
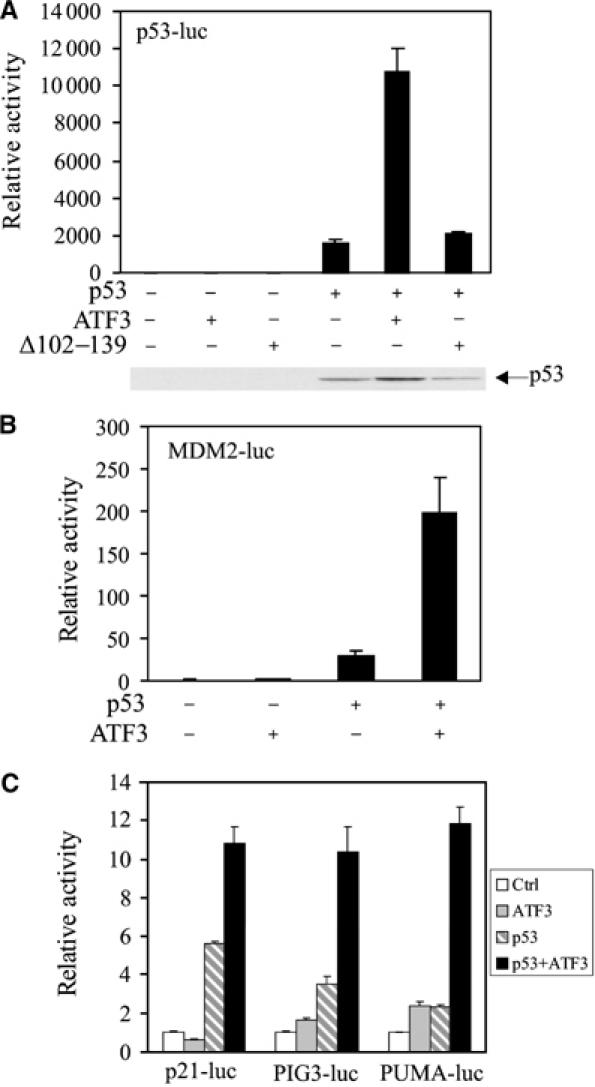
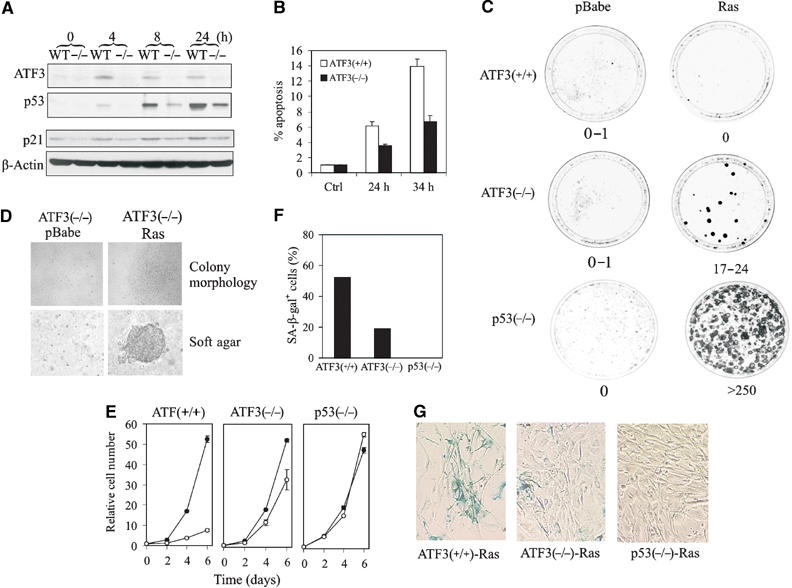
Activating transcription factor 3 (ATF3) is rapidly induced by diverse environmental insults including genotoxic stress. We report herein that its interaction with p53, enhanced by genotoxic stress, stabilizes the tumor suppressor thereby augmenting functions of the latter. Overexpression of ATF3 (but not a mutated ATF3 protein (Delta102-139) devoid of its p53-binding region) prevents p53 from MDM2-mediated degradation and leads to increased transcription from p53-regulated promoters. ATF3, but not the Delta102-139 protein, binds the p53 carboxy-terminus and diminishes its ubiquitination and nuclear export. Genotoxic-stressed ATF3-null mouse embryonic fibroblasts, or cells in which ATF3 was reduced by small interference RNA, show inefficient p53 induction and impaired apoptosis compared with wild-type cells. ATF3-null cells (but not wild-type cells), which poorly accumulate p53, are transformed by oncogenic Ras. Thus, ATF3 is a novel stress-activated regulator of p53 protein stability/function providing the cell with a means of responding to a wide range of environmental insult, thus maintaining DNA integrity and protecting against cell transformation.
Figures
Similar articles
-
YY1 inhibits the activation of the p53 tumor suppressor in response to genotoxic stress.Proc Natl Acad Sci U S A. 2004 Aug 17;101(33):12165-70. doi: 10.1073/pnas.0402283101. Epub 2004 Aug 4. Proc Natl Acad Sci U S A. 2004. PMID: 15295102 Free PMC article.
-
Competitive ubiquitination activates the tumor suppressor p53.Cell Death Differ. 2020 Jun;27(6):1807-1818. doi: 10.1038/s41418-019-0463-x. Epub 2019 Dec 2. Cell Death Differ. 2020. PMID: 31796886 Free PMC article.
-
p53 gene family p51(p63)-encoded, secondary transactivator p51B(TAp63alpha) occurs without forming an immunoprecipitable complex with MDM2, but responds to genotoxic stress by accumulation.Exp Cell Res. 2002 Jun 10;276(2):194-200. doi: 10.1006/excr.2002.5535. Exp Cell Res. 2002. PMID: 12027449
-
Control of p53 ubiquitination and nuclear export by MDM2 and ARF.Cell Growth Differ. 2001 Apr;12(4):175-86. Cell Growth Differ. 2001. PMID: 11331246 Review.
-
A new twist in the feedback loop: stress-activated MDM2 destabilization is required for p53 activation.Cell Cycle. 2005 Mar;4(3):411-7. doi: 10.4161/cc.4.3.1522. Epub 2005 Mar 2. Cell Cycle. 2005. PMID: 15684615 Review.
Cited by
-
Identification of ATF3 as a novel protective signature of quiescent colorectal tumor cells.Cell Death Dis. 2023 Oct 13;14(10):676. doi: 10.1038/s41419-023-06204-1. Cell Death Dis. 2023. PMID: 37833290 Free PMC article.
-
Locally acting transcription factors regulate p53-dependent cis-regulatory element activity.Nucleic Acids Res. 2020 May 7;48(8):4195-4213. doi: 10.1093/nar/gkaa147. Nucleic Acids Res. 2020. PMID: 32133495 Free PMC article.
-
Targeting HOX/PBX dimers in cancer.Oncotarget. 2017 May 9;8(19):32322-32331. doi: 10.18632/oncotarget.15971. Oncotarget. 2017. PMID: 28423659 Free PMC article. Review.
-
Impact of protein kinase PKR in cell biology: from antiviral to antiproliferative action.Microbiol Mol Biol Rev. 2006 Dec;70(4):1032-60. doi: 10.1128/MMBR.00027-06. Microbiol Mol Biol Rev. 2006. PMID: 17158706 Free PMC article. Review.
-
A small-molecule inhibitor of MDMX activates p53 and induces apoptosis.Mol Cancer Ther. 2011 Jan;10(1):69-79. doi: 10.1158/1535-7163.MCT-10-0581. Epub 2010 Nov 12. Mol Cancer Ther. 2011. PMID: 21075910 Free PMC article.
References
-
- An WG, Kanekal M, Aaronswon SA, Simon MC, Maltepe E, Blagosklonny MV, Neckers LM (1998) Stabilization of wild-type p53 by hypoxia-inducible factor 1α. Nature 392: 405–408 - PubMed
-
- Buschmann T, Lin Y, Aithmitti N, Fuchs SY, Lu H, Resnick-Silverman L, Manfredi JJ, Ronai Z, Wu X (2001) Stabilization and activation of p53 by the coactivator protein TAFII31. J Biol Chem 276: 13852–13857 - PubMed
-
- Chen D, Li M, Luo J, Gu W (2003) Direct interactions between HIF-1α and Mdm2 modulate p53 function. J Biol Chem 278: 13595–13598 - PubMed
-
- Cheng T, Liu D, Griffin JH, Fernandez JA, Castellino F, Rosen ED, Fukudome K, Zlokovic BV (2003) Activated protein C blocks p53-mediated apoptosis in ischemic human brain endothelium and is neuroprotective. Nat Med 9: 338–342 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
