

PIASy mediates SUMO-2 conjugation of Topoisomerase-II on mitotic chromosomes
- PMID: 15933717
- PMCID: PMC1150894
- DOI: 10.1038/sj.emboj.7600700
PIASy mediates SUMO-2 conjugation of Topoisomerase-II on mitotic chromosomes
Abstract
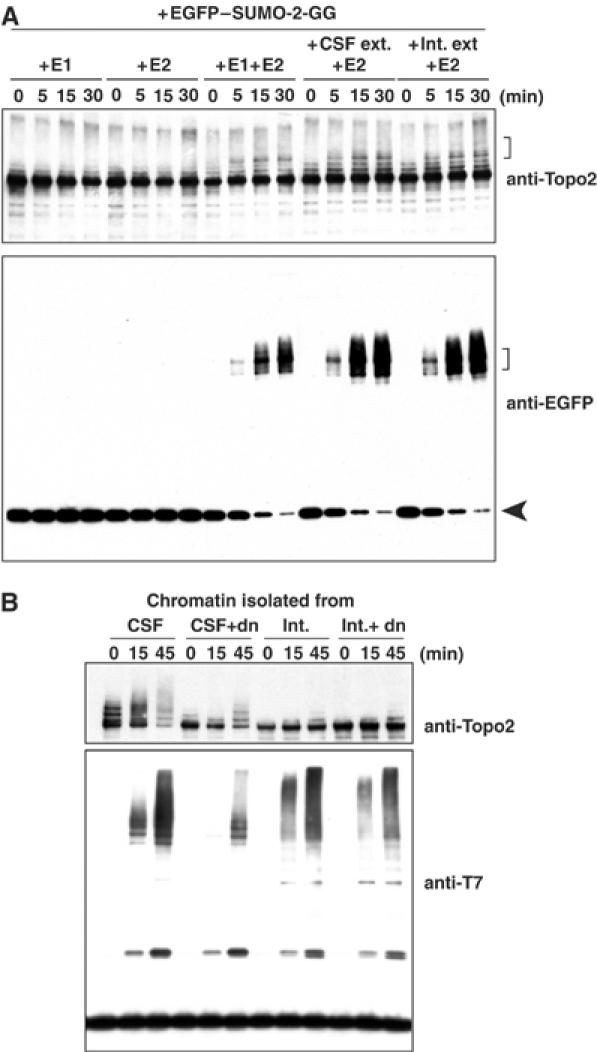
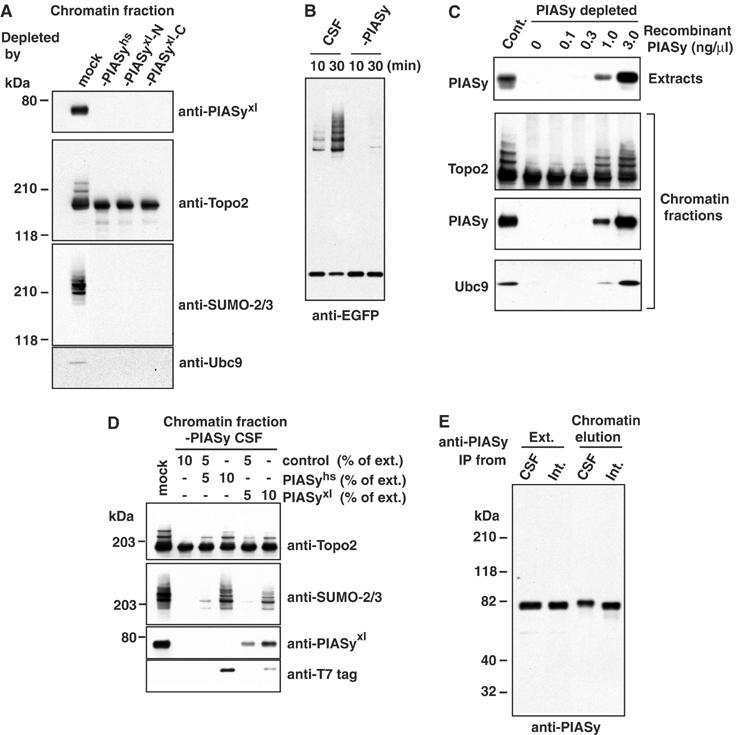
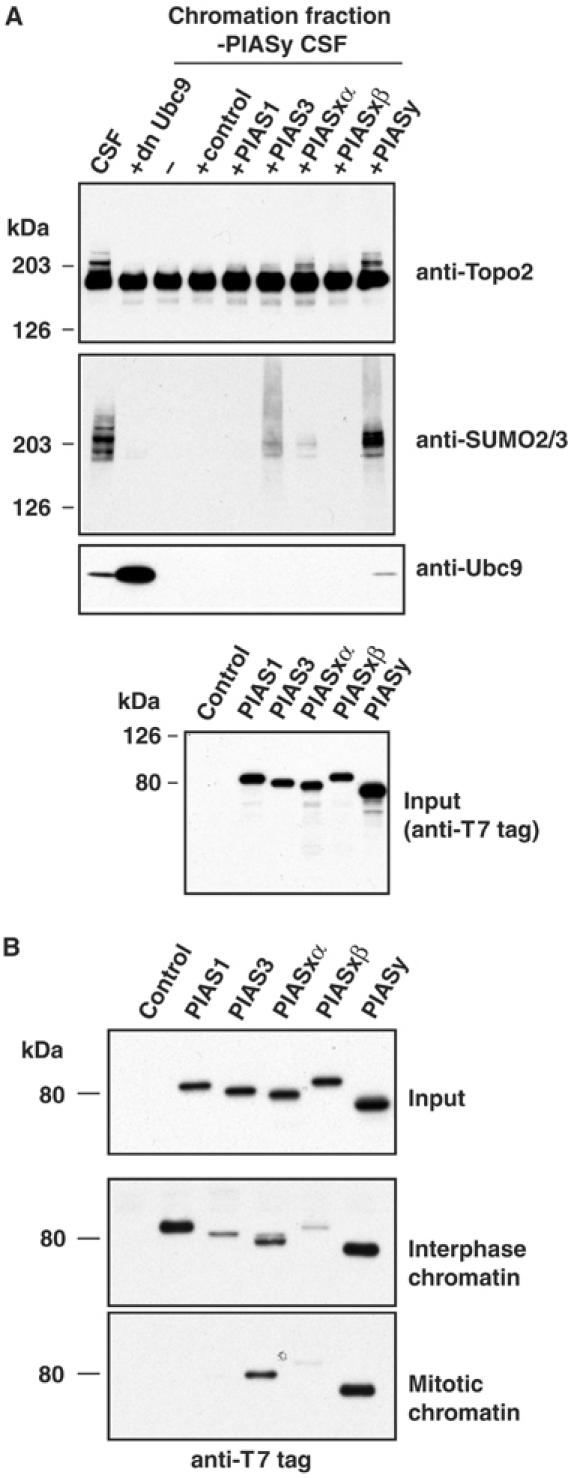
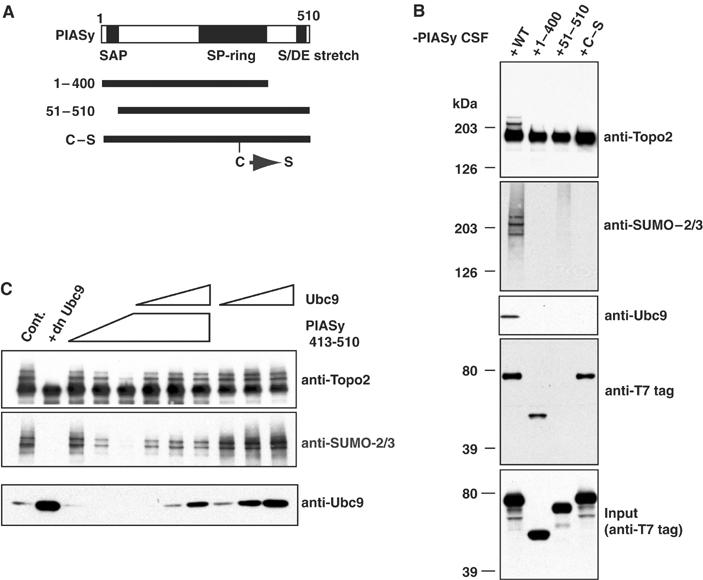
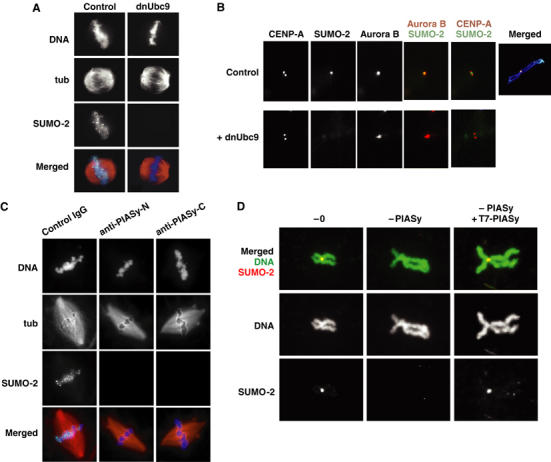
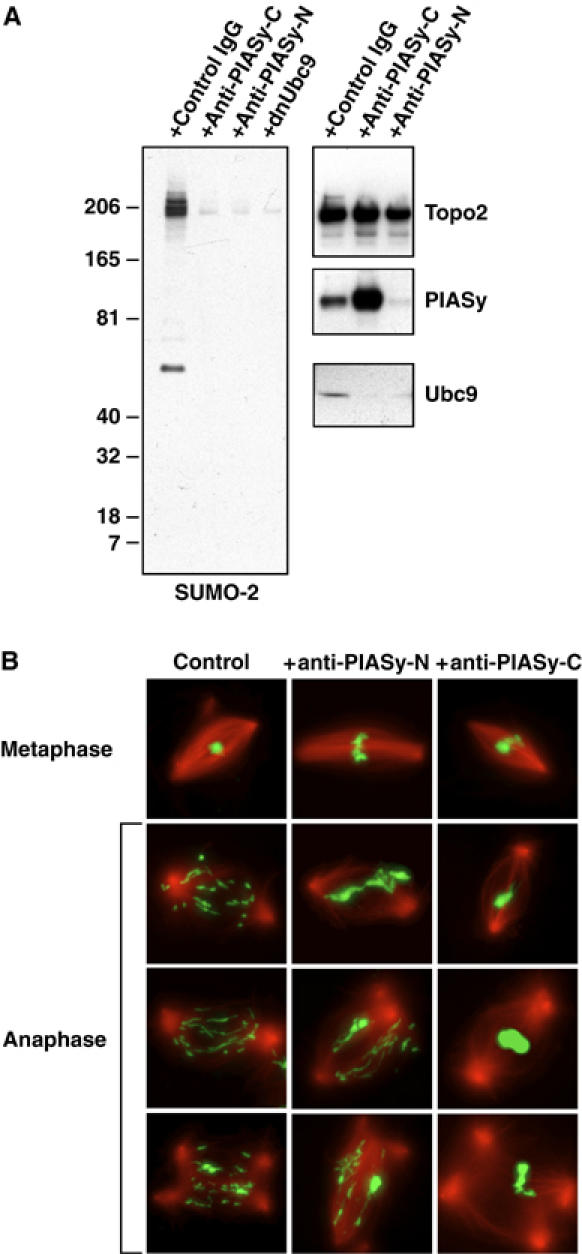
Here we show that the PIASy protein is specifically required for mitotic modification of Topoisomerase-II by SUMO-2 conjugation in Xenopus egg extracts. PIASy was unique among the PIAS family members in its capacity to bind mitotic chromosomes and recruit Ubc9 onto chromatin. These properties were essential, since PIASy mutants that did not bind chromatin or failed to recruit Ubc9 were functionally inactive. We observed that PIASy depletion eliminated essentially all chromosomal accumulation of EGFP-SUMO-2-conjugated species, suggesting that it is the primary E3-like factor for mitotic chromosomal substrates of SUMO-2. PIASy-dependent SUMO-2-conjugated species were highly concentrated on the inner centromere, and inhibition of PIASy blocked anaphase sister chromatid segregation in egg extracts. Taken together, our observations suggest that PIASy is a critical regulator of mitotic SUMO-2 conjugation for Topoisomerase-II and other chromosomal substrates, and that its activity may have particular relevance for centromeric functions required for proper chromosome segregation.
Figures
References
-
- Aravind L, Koonin EV (2000) SAP—a putative DNA-binding motif involved in chromosomal organization. Trends Biochem Sci 25: 112–114 - PubMed
-
- Arnaoutov A, Dasso M (2003) The Ran GTPase regulates kinetochore function. Dev Cell 5: 99–111 - PubMed
-
- Azuma Y, Tan SH, Cavenagh MM, Ainsztein AM, Saitoh H, Dasso M (2001) Expression and regulation of the mammalian SUMO-1 E1 enzyme. FASEB J 15: 1825–1827 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
