

Formation of the BMP activity gradient in the Drosophila embryo
- PMID: 15935780
- PMCID: PMC1829168
- DOI: 10.1016/j.devcel.2005.04.009
Formation of the BMP activity gradient in the Drosophila embryo
Abstract
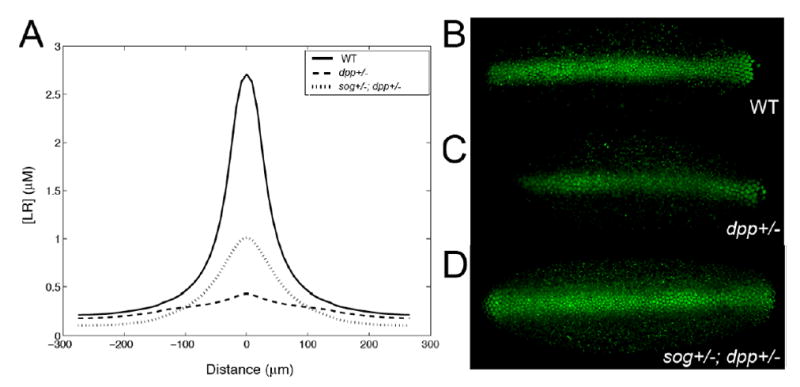
The dorsoventral axis of the Drosophila embryo is patterned by a gradient of bone morphogenetic protein (BMP) ligands. In a process requiring at least three additional extracellular proteins, a broad domain of weak signaling forms and then abruptly sharpens into a narrow dorsal midline peak. Using experimental and computational approaches, we investigate how the interactions of a multiprotein network create the unusual shape and dynamics of formation of this gradient. Starting from observations suggesting that receptor-mediated BMP degradation is an important driving force in gradient dynamics, we develop a general model that is capable of capturing both subtle aspects of gradient behavior and a level of robustness that agrees with in vivo results.
Figures

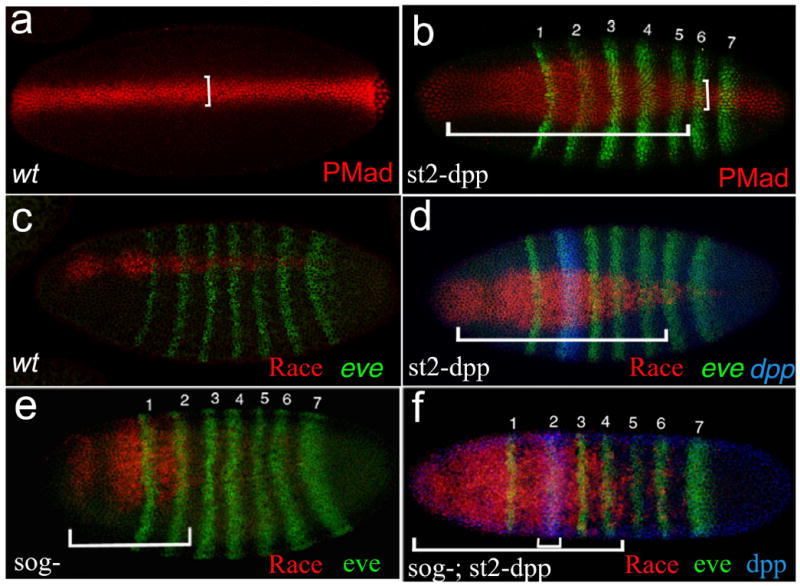

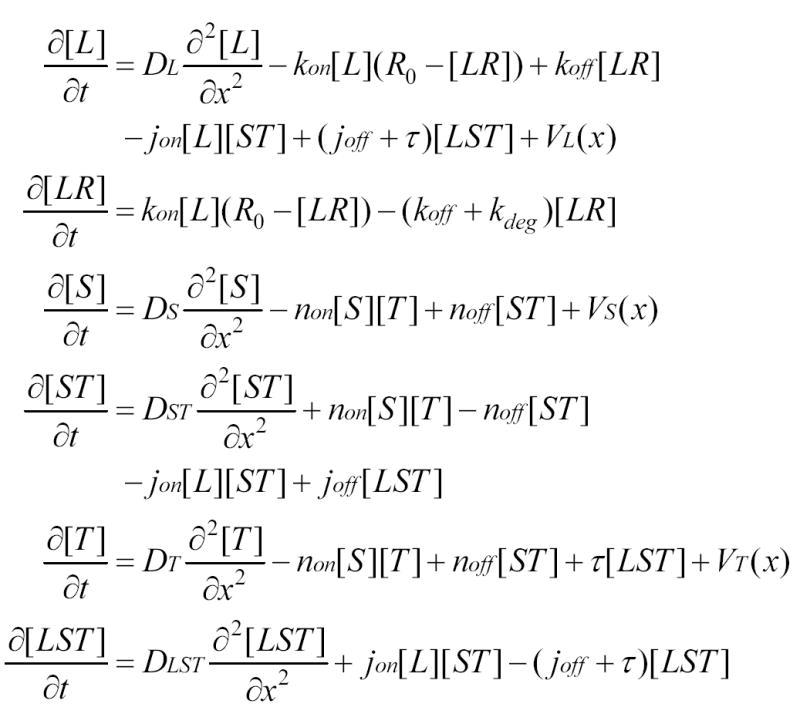
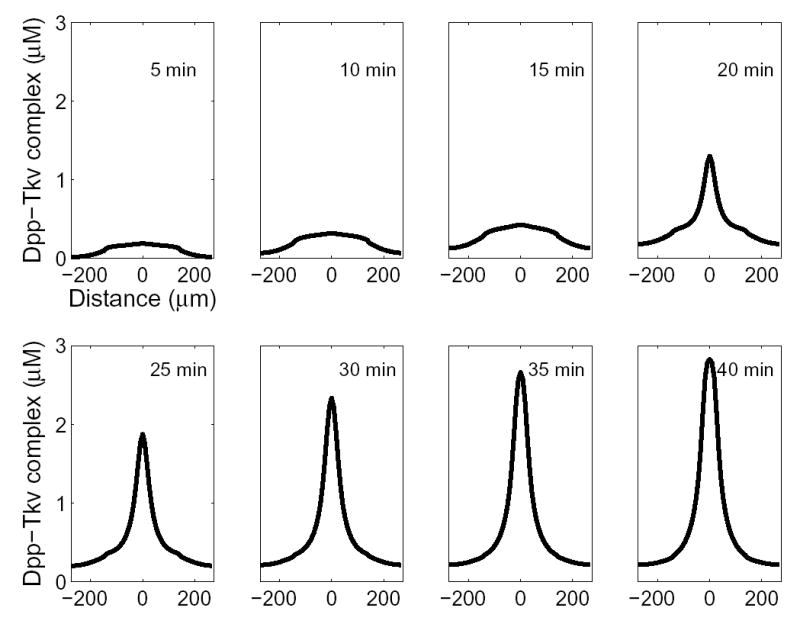
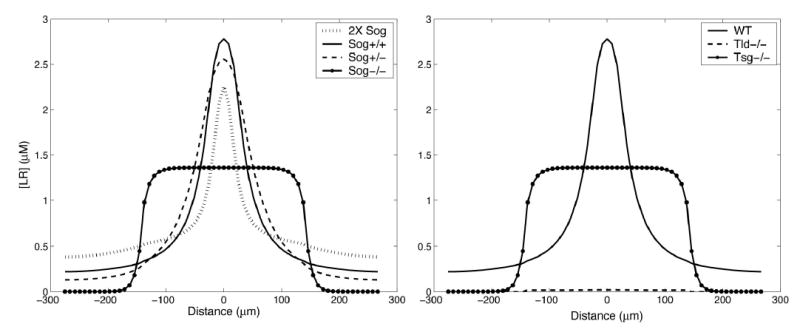
References
-
- Alon U, Surette MG, Barkai N, Leibler S. Robustness in bacterial chemotaxis. Nature. 1999;397:168–171. - PubMed
-
- Arora K, Levine MS, O’Connor MB. The screw gene encodes a ubiquitously expressed member of the TGF-beta family required for specification of dorsal cell fates in the Drosophila embryo. Genes Dev. 1994;8:2588–2601. - PubMed
-
- Ashe HL. BMP signalling: visualisation of the Sog protein gradient. Curr Biol. 2002;12:R273–R275. - PubMed
-
- Ashe HL, Levine M. Local inhibition and long-range enhancement of Dpp signal transduction by Sog. Nature. 1999;398:427–431. - PubMed
-
- Ashe HL, Mannervik M, Levine M. Dpp signaling thresholds in the dorsal ectoderm of the Drosophila embryo. Development. 2000;127:3305–3312. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
