

Review
doi: 10.1098/rstb.2005.1624.
The future of mapping sensory cortex in primates: three of many remaining issues
Affiliations
- PMID: 15937006
- PMCID: PMC1569483
- DOI: 10.1098/rstb.2005.1624
Item in Clipboard
Review
The future of mapping sensory cortex in primates: three of many remaining issues
Philos Trans R Soc Lond B Biol Sci.
.
Abstract
After 100 years of progress in understanding the organization of cerebral cortex, three issues have persisted over the last 35 years, which are revisited in this paper. First, is V3 an established or questionable area of visual cortex? Second, does taste cortex include part of area 3b (S1 proper) and other somatosensory areas? Third, is primary auditory cortex, A1, of primates the homologue of A1 in cats? The existence of such questions about even the early stages of cortical processing reflects the difficulties in mapping cerebral cortex, and reminds us that the era of basic discovery is far from over.
Figures

Different proposals for how visual cortex in the region of V3 is organized. (a) Traditional proposals stemming from the descriptions of Cragg (1969) and Zeki (1969) include a V3 with a dorsal half, V3d, representing the lower visual quadrant, and a ventral half, V3v, representing the upper visual quadrant. The second visual area, V2, is split along the zero horizontal meridian (HM) to border primary visual cortex, V1, along the representation of the zero vertical meridian (VM). Area V3 borders much of V2, roughly as a mirror image of V2. V1 projects to retinotopically matched sets in both V2 and V3, as well as other areas including the dorsomedial visual area, DM, possibly equivalent to V3A. Sites along the outer border of V1 correspond to the VM and project to the outer border of V3, while sites along the midline of V1 correspond to the HM and project to the V2/V3 border. In the original portrayals of V3 of Cragg (1969) and Zeki (1969), V2 and V3 were continuous and of equal size. It now appears likely that in at least some primates, V3 has separated dorsal (V3d) and ventral (V3v) halves, and V3 is smaller than V2 (e.g. Kaas & Lyon 2001). (b) An alternative proposal is that a number of visual areas border V2, and there is no V3 (Allman & Kaas 1974). In this view, much of V3d is subsumed in DM, and projections of V1 that were considered to be to V3d are instead to DM (Lin et al., 1982). Because the evidence for V1 projections to V3v was uncertain, the region of V3v was considered part of another visual area (a combination of ventroposterior, VP, and ventroanterior, VA, visual areas (see Newsome & Allman 1980). (c) Perhaps the dominant theory today is that V3d is a visual area by itself even though it represents only the lower visual quadrant. The V3v region is also considered a separate visual area, the ventroposterior area, VP, which represents only the upper visual quadrant. This theory (as well as those in (b) and (d)) was supported by the erroneous conclusion that V3v does not have connections with V1 (Weller & Kaas 1983; Van Essen et al. 1986). (d) Another possibility is that much of V3d is part of DM (as in (b) above), while V3v is part of a larger area, the ventrolateral posterior area (VLP). The dorsolateral complex (DL or V4) of monkeys has been divided into rostral (DLr) and caudal (DLc) areas (see Stepniewska & Kaas 1996) and DLc has been combined with V3v/VP to form VLP by Rosa & Tweedale (2000). They have also combined the territory of DLr with the region of VA or V4v to form the ventrolateral anterior area, VLA.
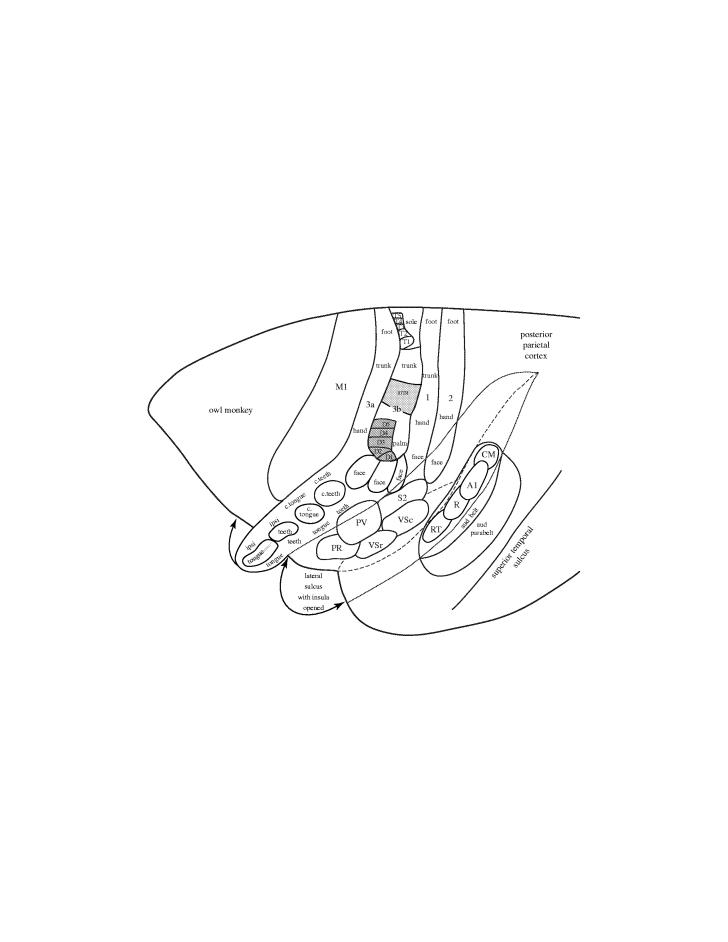
A lateral view of the rostral two-thirds of the left cerebral hemisphere of an owl monkey. The full extent of primary somatosensory cortex (area 3b) in New World owl and squirrel monkeys, as recently defined by Jain et al. (2001), includes a large RL representation of the oral cavity with representations of the periodontal receptors of the contralateral teeth (c. teeth), the contralateral tongue (c. tongue), ipsilateral teeth (ipsi teeth) and ipsilateral tongue (ipsi tongue) in a dorsocaudal to ventrorostral sequence. In the more medial area 3b, three ovals of cortex represent the lower face and lower lip, the upper lip and the upper face. These ovals are followed by representations of the hand with digits (1–5), arm, trunk and sole and toes of the foot. Area 3b is bordered by rostral and caudal parallel representations of the contralateral body in areas 3a and 1. These areas appear to extend rostrolaterally to form representations of the oral cavity that adjoin that of area 3b. Additional somatosensory representations include the second somatosensory area, S2, the parietal ventral area, PV, the rostral ventral somatosensory area, VSr, the caudal ventral somatosensory area, VSc, and the parietal rostral area, PR (see Coq et al. 2004). The lateral sulcus has been opened to show these areas and the insular cortex. Area 2 contains another representation of the contralateral body. Many or most of these somatosensory areas represent the tongue, and some of them receive direct inputs from the taste nucleus of the thalamus. Thus, tongue portions of all or most of these areas, in addition to other fields in the orbital–frontal cortex, may be involved in a complex cortical network that mediates aspects of taste. Primary motor cortex, M1 and auditory areas are shown for reference. Primary auditory cortex, A1, the primary-like rostral area, R, and rostral temporal area, RT. The auditory (aud) belt, of secondary auditory areas includes the caudomedial area, CM. The auditory (aud) parabelt constitutes a third level of cortical processing.

The tonotopic organization of three primary-like auditory fields in (a) cats (and other carnivores) and monkeys (b). Cats have an anterior auditory field, AAF, a primary field, A1, and a posterior field, P. These fields represent high to low frequencies in isofrequency bands that progress from rostral to caudal in A1, but in the opposite (mirror-image) patterns in the two adjoining fields. Monkeys have a rostral–temporal field, RT, a rostral field, R, and a primary field, A1. The rostrocaudal tonotopic sequence of representation in R matches that of A1 of cats, except for a tilt, while RT and A1 mirror R in tonotopic organization. The question mark (?) for RT denotes the sparseness of data on the tonotopy of the area; R, rostral in the brain; C, caudal.
Similar articles
-
[Multiple sensory projections in the dolphin cerebral cortex].Zh Vyssh Nerv Deiat Im I P Pavlova. 1978 Sep-Oct;28(5):1047-53. Zh Vyssh Nerv Deiat Im I P Pavlova. 1978. PMID: 716593 Russian.
-
[Localization of the sensory projection areas in the cerebral cortex of the dolphin, Tursiops truncatus].Zh Evol Biokhim Fiziol. 1977 Nov-Dec;13(6):712-8. Zh Evol Biokhim Fiziol. 1977. PMID: 602532 Russian.
-
Auditory processing in primate cerebral cortex.Curr Opin Neurobiol. 1999 Apr;9(2):164-70. doi: 10.1016/s0959-4388(99)80022-1. Curr Opin Neurobiol. 1999. PMID: 10322185 Review.
-
Hearing taste and colouring text.Lancet Neurol. 2005 May;4(5):274. doi: 10.1016/s1474-4422(05)70062-4. Lancet Neurol. 2005. PMID: 15861554 No abstract available.
-
Auditory processing and hemispheric specialization in non-human primates.Cortex. 2006 Jan;42(1):87-9. doi: 10.1016/s0010-9452(08)70325-3. Cortex. 2006. PMID: 16509112 Review. No abstract available.
Cited by
-
Increased Subjective Distaste and Altered Insula Activity to Umami Tastant in Patients with Bulimia Nervosa.Front Psychiatry. 2017 Sep 25;8:172. doi: 10.3389/fpsyt.2017.00172. eCollection 2017. Front Psychiatry. 2017. PMID: 28993739 Free PMC article.
-
A Web-Based Atlas Combining MRI and Histology of the Squirrel Monkey Brain.Neuroinformatics. 2019 Jan;17(1):131-145. doi: 10.1007/s12021-018-9391-z. Neuroinformatics. 2019. PMID: 30006920 Free PMC article.
-
Introduction: cerebral cartography 1905-2005.Philos Trans R Soc Lond B Biol Sci. 2005 Apr 29;360(1456):651-2. doi: 10.1098/rstb.2005.1632. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15937005 Free PMC article. No abstract available.
-
The representation of oral fat texture in the human somatosensory cortex.Hum Brain Mapp. 2014 Jun;35(6):2521-30. doi: 10.1002/hbm.22346. Epub 2013 Sep 3. Hum Brain Mapp. 2014. PMID: 24038614 Free PMC article.
-
Interconnected sub-networks of the macaque monkey gustatory connectome.Front Neurosci. 2023 Feb 16;16:818800. doi: 10.3389/fnins.2022.818800. eCollection 2022. Front Neurosci. 2023. PMID: 36874640 Free PMC article.
References
-
- Allman J.M, Kaas J.H. Representation of the visual field in striate and adjoining cortex of the owl monkey (Aotus trivirgatus) Brain Res. 1971;35:89–106. - PubMed
-
- Allman J.M, Kaas J.H. The organization of the second visual area (VII) in the owl monkey: a second order transformation of the visual hemifield. Brain Res. 1974;76:247–265. - PubMed
-
- Allman J.M, Kaas J.H. Representation of the visual field on the medial wall of occipital–parietal cortex in the owl monkey. Science. 1976;191:572–575. - PubMed
-
- Beck P.D, Kaas J.H. Cortical connections of the dorsomedial visual area in old world macaque monkeys. J. Comp. Neurol. 1999;406:487–502. - PubMed
-
- Benjamin R.M, Burton H. Projection of taste nerve afferents to anterior opercular–insular cortex in squirrel monkey (Saimiri sciureus) Brain Res. 1968;7:221–231. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
