

Small-molecule inhibition of proteasome and aggresome function induces synergistic antitumor activity in multiple myeloma
- PMID: 15937109
- PMCID: PMC1150844
- DOI: 10.1073/pnas.0503221102
Small-molecule inhibition of proteasome and aggresome function induces synergistic antitumor activity in multiple myeloma
Abstract
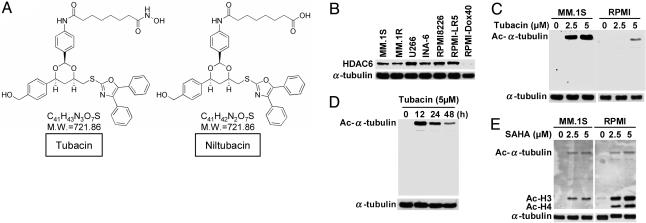
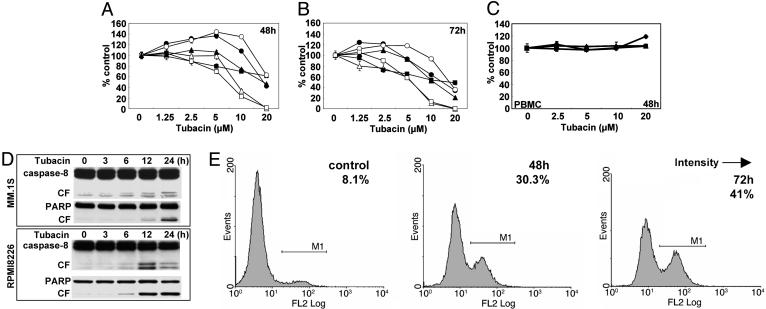
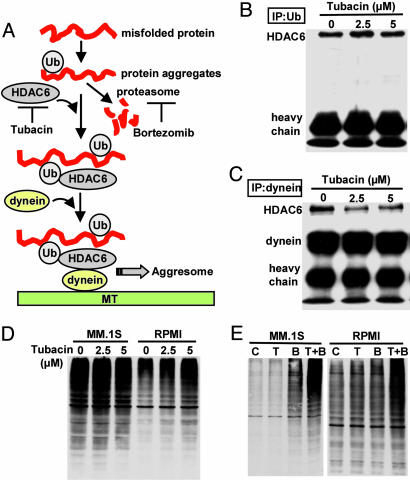
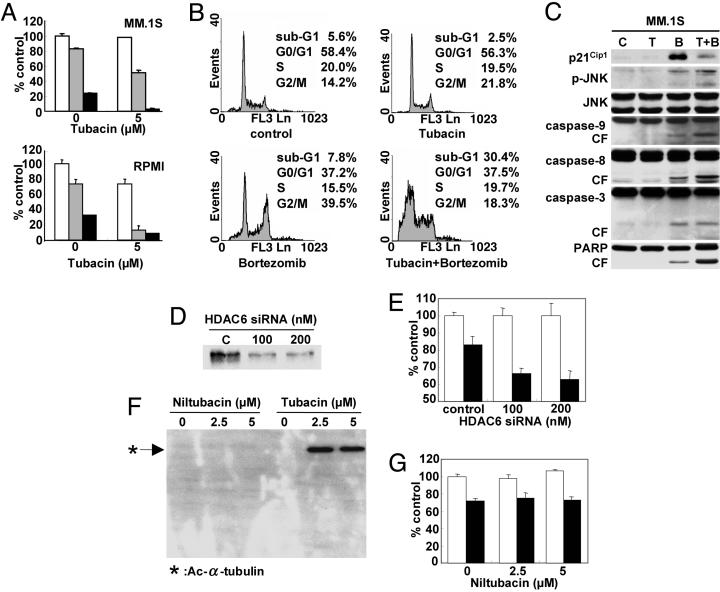
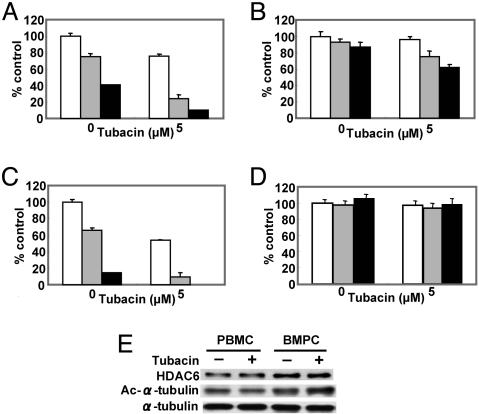
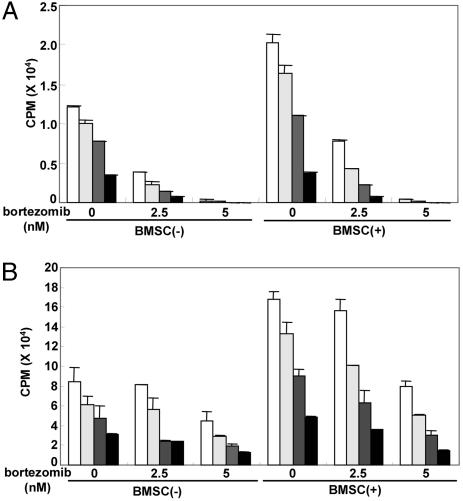
We have shown that the proteasome inhibitor bortezomib (formerly known as PS-341) triggers significant antitumor activity in multiple myeloma (MM) in both preclinical models and patients with relapsed refractory disease. Recent studies have shown that unfolded and misfolded ubiquitinated proteins are degraded not only by proteasomes, but also by aggresomes, dependent on histone deacetylase 6 (HDAC6) activity. We therefore hypothesized that inhibition of both mechanisms of protein catabolism could induce accumulation of ubiquitinated proteins followed by significant cell stress and cytotoxicity in MM cells. To prove this hypothesis, we used bortezomib and tubacin to inhibit the proteasome and HDAC6, respectively. Tubacin specifically triggers acetylation of alpha-tubulin as a result of HDAC6 inhibition in a dose- and time-dependent fashion. It induces cytotoxicity in MM cells at 72 h with an IC50 of 5-20 microM, which is mediated by caspase-dependent apoptosis; no toxicity is observed in normal peripheral blood mononuclear cells. Tubacin inhibits the interaction of HDAC6 with dynein and induces marked accumulation of ubiquitinated proteins. It synergistically augments bortezomib-induced cytotoxicity by c-Jun NH2-terminal kinase/caspase activation. Importantly, this combination also induces significant cytotoxicity in plasma cells isolated from MM patient bone marrow. Finally, adherence of MM cells to bone marrow stromal cells confers growth and resistance to conventional treatments; in contrast, the combination of tubacin and bortezomib triggers toxicity even in adherent MM cells. Our studies therefore demonstrate that tubacin combined with bortezomib mediates significant anti-MM activity, providing the framework for clinical evaluation of combined therapy to improve patient outcome in MM.
Figures
References
-
- Gregory, W. M., Richards, M. A. & Malpas, J. S. (1992) J. Clin. Oncol. 10, 334-342. - PubMed
-
- Attal, M., Harousseau, J. L., Facon, T., Guilhot, F., Doyen, C., Fuzibet, J. G., Monconduit, M., Hulin, C., Caillot, D., Bouabdallah, R., et al. (2003) N. Engl. J. Med. 349, 2495-2502. - PubMed
-
- Hideshima, T. & Anderson, K. C. (2002) Nat. Rev. Cancer 2, 927-937. - PubMed
-
- Hideshima, T., Richardson, P., Chauhan, D., Palombella, V., Elliott, P., Adams, J. & Anderson, K. C. (2001) Cancer Res. 61, 3071-3076. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
