

Cell-type-specific epigenetic marking of the IL2 gene at a distal cis-regulatory region in competent, nontranscribing T-cells
- PMID: 15937196
- PMCID: PMC1142491
- DOI: 10.1093/nar/gki637
Cell-type-specific epigenetic marking of the IL2 gene at a distal cis-regulatory region in competent, nontranscribing T-cells
Abstract
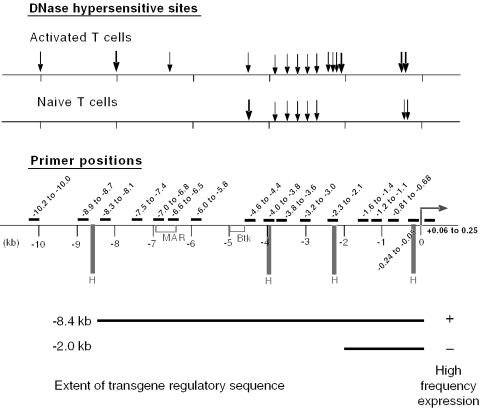
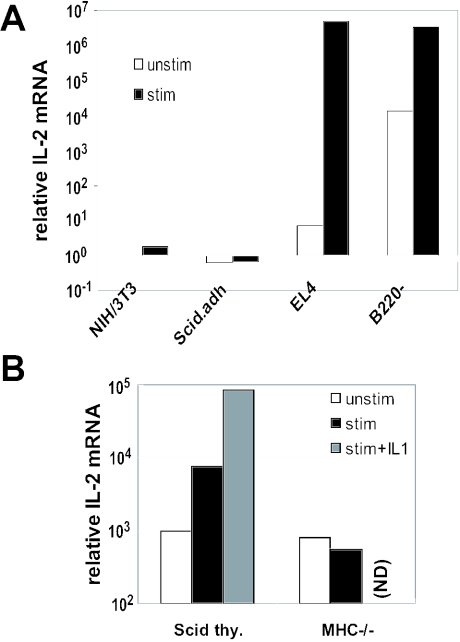
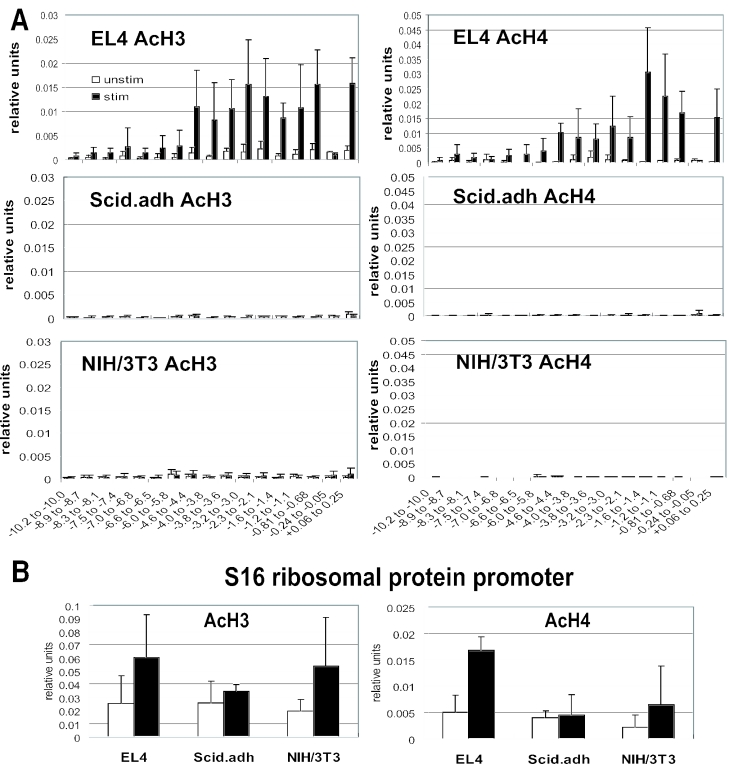
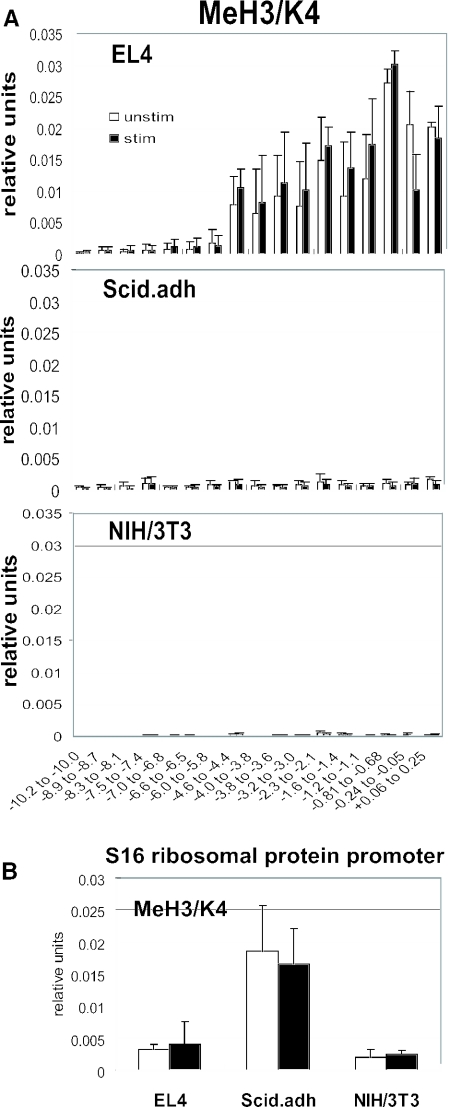
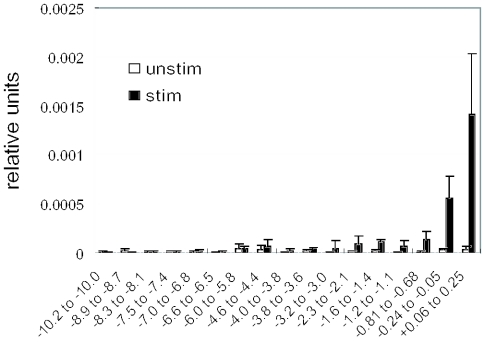
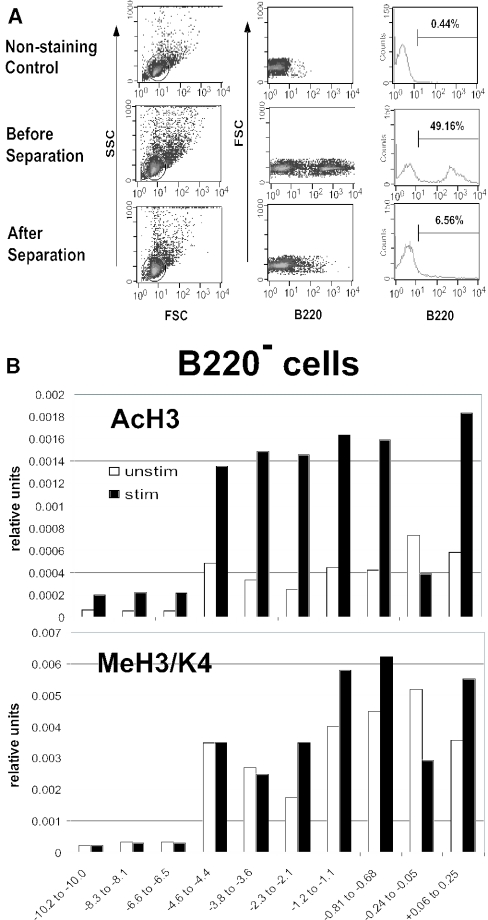
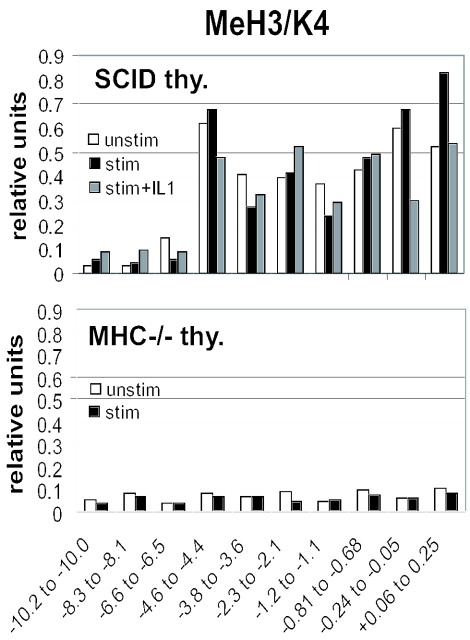
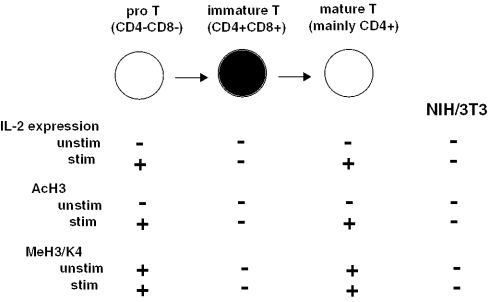
T-cells retain cell-type-specific programming for IL-2 inducibility through many rounds of division without being stimulated to transcribe the locus. To understand the layering of controls needed to poise this gene heritably for activation, we have used chromatin immunoprecipitation to map histone modifications across the murine IL2 locus, from -10.2 through +0.25 kb, in induction-competent and incompetent cells. In highly inducible EL4 T-lineage cells, stimulation with PMA/A23187 induced strong acetylation of histone H3 and H4, in parallel with transcriptional activation, from -4.6 through +0.25 kb. However, dimethylation of histone H3/K4 was already fully elevated across the same restricted domain before stimulation, with little change after stimulation. RNA polymerase II binding, in contrast, was only found at the known promoter region after stimulation. Similar patterns of histone modifications were seen also in normal IL-2-inducible T-lineage cells. However, neither acetylated histone H3, H4 nor dimethylated histone H3/K4 marking was detected, with or without stimulation, in expression-incompetent cells (NIH/3T3 or Scid.adh). These results identify a discrete new domain of IL2 regulatory sequence marked by dimethylated histone H3/K4 in expression-permissive T-cells even when they are not transcribing IL2, setting boundaries for histone H3 and H4 acetylation when the IL2 gene is transcriptionally activated.
Figures
References
-
- Reya T., Morrison S.J., Clarke M.F., Weissman I.L. Stem cells, cancer, and cancer stem cells. Nature. 2001;414:105–111. - PubMed
-
- Fischle W., Wang Y., Allis C.D. Binary switches and modification cassettes in histone biology and beyond. Nature. 2003;425:475–479. - PubMed
-
- Marmorstein R. Protein modules that manipulate histone tails for chromatin regulation. Nature Rev. Mol. Cell Biol. 2001;2:422–432. - PubMed
-
- Yamashita M., Ukai-Tadenuma M., Miyamoto T., Sugaya K., Hosokawa H., Hasegawa A., Kimura M., Taniguchi M., DeGregori J., Nakayama T. Essential role of GATA3 for the maintenance of type 2 helper T (Th2) cytokine production and chromatin remodeling at the Th2 cytokine gene loci. J. Biol. Chem. 2004;279:26983–26990. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
