

Systemic acquired tolerance to virulent bacterial pathogens in tomato
- PMID: 15937273
- PMCID: PMC1176419
- DOI: 10.1104/pp.105.059246
Systemic acquired tolerance to virulent bacterial pathogens in tomato
Abstract
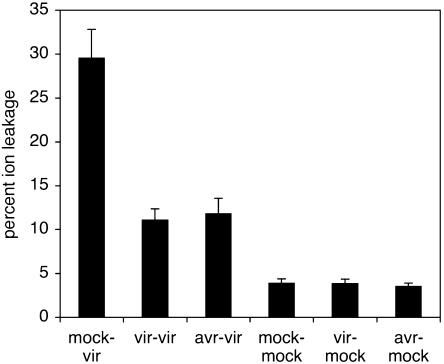
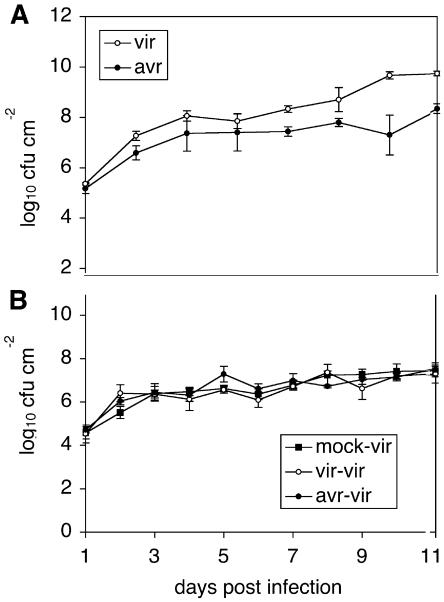
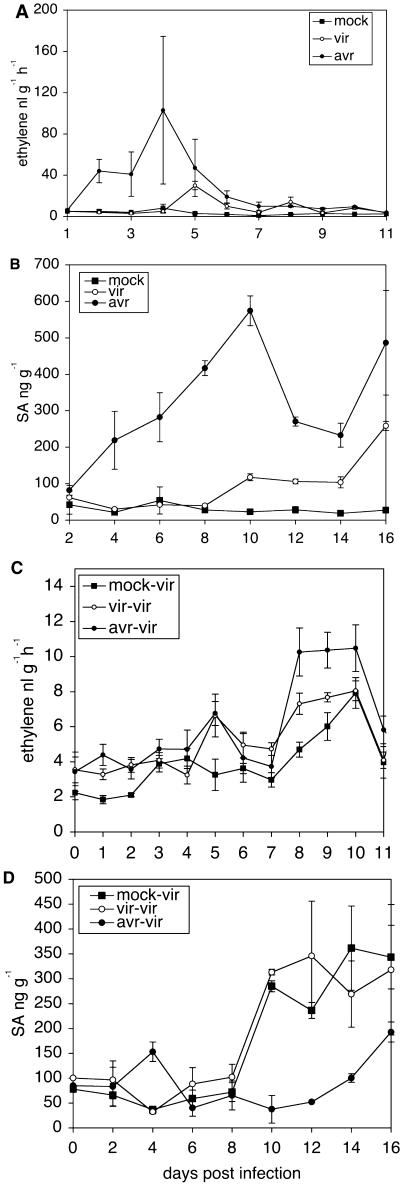
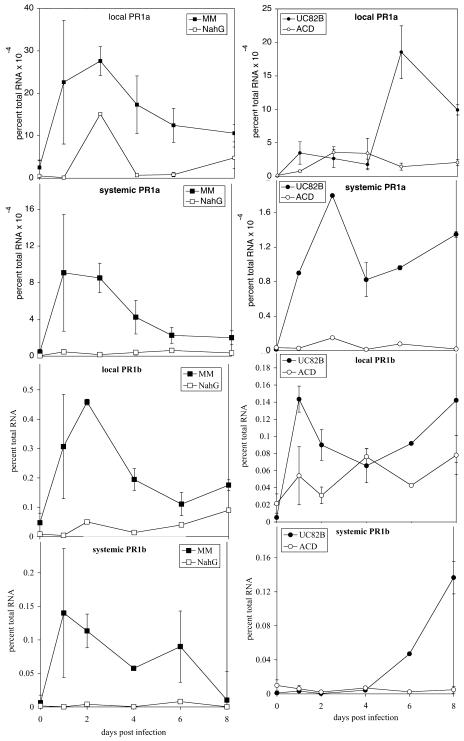
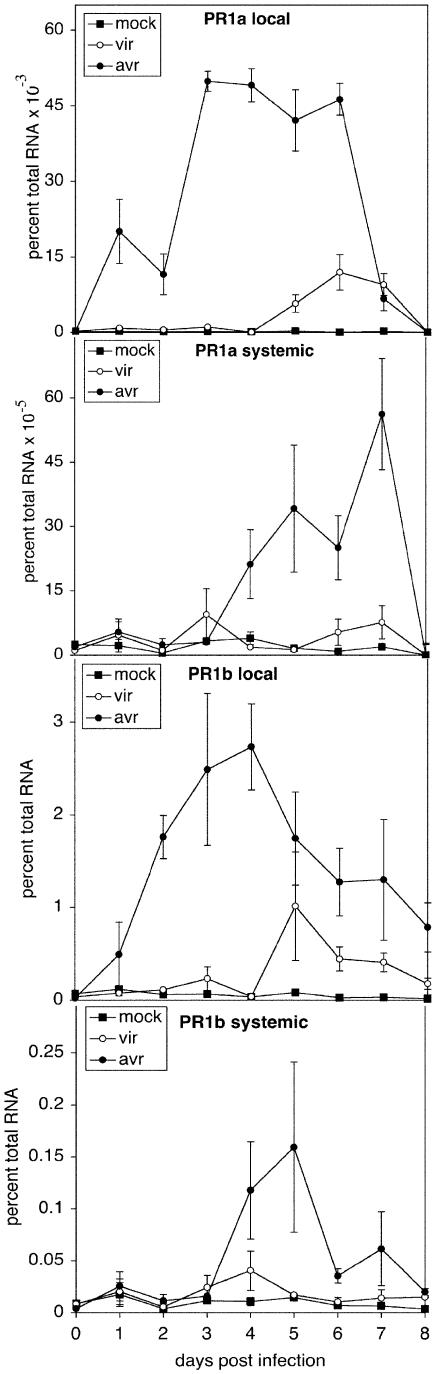
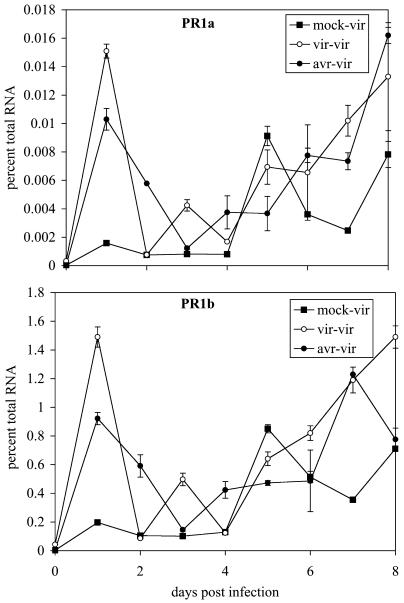
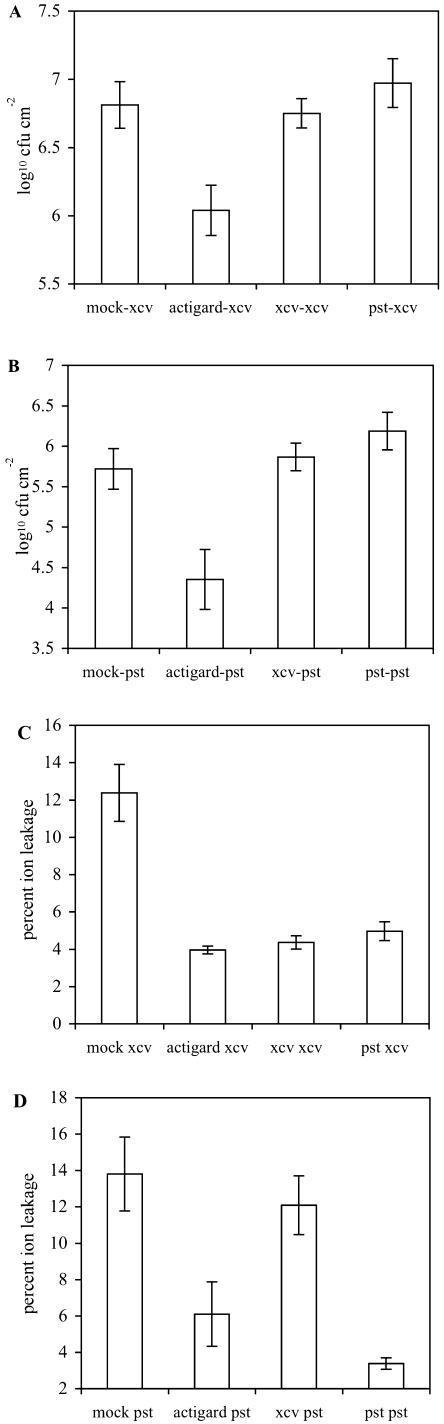
Recent studies on the interactions between plants and pathogenic microorganisms indicate that the processes of disease symptom development and pathogen growth can be uncoupled. Thus, in many instances, the symptoms associated with disease represent an active host response to the presence of a pathogen. These host responses are frequently mediated by phytohormones. For example, ethylene and salicylic acid (SA) mediate symptom development but do not influence bacterial growth in the interaction between tomato (Lycopersicon esculentum) and virulent Xanthomonas campestris pv vesicatoria (Xcv). It is not apparent why extensive tissue death is integral to a defense response if it does not have the effect of limiting pathogen proliferation. One possible function for this hormone-mediated response is to induce a systemic defense response. We therefore assessed the systemic responses of tomato to Xcv. SA- and ethylene-deficient transgenic lines were used to investigate the roles of these phytohormones in systemic signaling. Virulent and avirulent Xcv did induce a systemic response as evidenced by expression of defense-associated pathogenesis-related genes in an ethylene- and SA-dependent manner. This systemic response reduced cell death but not bacterial growth during subsequent challenge with virulent Xcv. This systemic acquired tolerance (SAT) consists of reduced tissue damage in response to secondary challenge with a virulent pathogen with no effect upon pathogen growth. SAT was associated with a rapid ethylene and pathogenesis-related gene induction upon challenge. SAT was also induced by infection with Pseudomonas syringae pv tomato. These data show that SAT resembles systemic acquired resistance without inhibition of pathogen growth.
Figures

Similar articles
-
Multiple hormones act sequentially to mediate a susceptible tomato pathogen defense response.Plant Physiol. 2003 Nov;133(3):1181-9. doi: 10.1104/pp.103.030379. Epub 2003 Oct 9. Plant Physiol. 2003. PMID: 14551324 Free PMC article.
-
Real time expression of ACC oxidase and PR-protein genes mediated by Methylobacterium spp. in tomato plants challenged with Xanthomonas campestris pv. vesicatoria.J Plant Physiol. 2014 Jul 15;171(12):1064-75. doi: 10.1016/j.jplph.2014.03.009. Epub 2014 Apr 4. J Plant Physiol. 2014. PMID: 24974333
-
The Pseudomonas syringae avrRpt2 gene contributes to virulence on tomato.Mol Plant Microbe Interact. 2005 Jul;18(7):626-33. doi: 10.1094/MPMI-18-0626. Mol Plant Microbe Interact. 2005. PMID: 16042008
-
Ethylene regulates the susceptible response to pathogen infection in tomato.Plant Cell. 1998 Mar;10(3):371-82. doi: 10.1105/tpc.10.3.371. Plant Cell. 1998. PMID: 9501111 Free PMC article.
-
Identification and expression profiling of tomato genes differentially regulated during a resistance response to Xanthomonas campestris pv. vesicatoria.Mol Plant Microbe Interact. 2004 Nov;17(11):1212-22. doi: 10.1094/MPMI.2004.17.11.1212. Mol Plant Microbe Interact. 2004. PMID: 15553246
Cited by
-
S-glycoprotein-like protein regulates defense responses in Nicotiana plants against Ralstonia solanacearum.Plant Physiol. 2010 Apr;152(4):2023-35. doi: 10.1104/pp.109.148189. Epub 2010 Jan 29. Plant Physiol. 2010. PMID: 20118275 Free PMC article.
-
Involvement of the pepper antimicrobial protein CaAMP1 gene in broad spectrum disease resistance.Plant Physiol. 2008 Oct;148(2):1004-20. doi: 10.1104/pp.108.123836. Epub 2008 Aug 1. Plant Physiol. 2008. PMID: 18676663 Free PMC article.
-
Ralstonia solanacearum requires PopS, an ancient AvrE-family effector, for virulence and To overcome salicylic acid-mediated defenses during tomato pathogenesis.mBio. 2013 Nov 26;4(6):e00875-13. doi: 10.1128/mBio.00875-13. mBio. 2013. PMID: 24281716 Free PMC article.
-
Effect of a benzothiadiazole on inducing resistance of soybean to Phytophthora sojae.Protoplasma. 2013 Apr;250(2):471-81. doi: 10.1007/s00709-012-0430-6. Epub 2012 Jul 10. Protoplasma. 2013. PMID: 22777214
-
Application of proteomics to investigate stress-induced proteins for improvement in crop protection.Plant Cell Rep. 2011 May;30(5):745-63. doi: 10.1007/s00299-010-0982-x. Epub 2011 Feb 2. Plant Cell Rep. 2011. PMID: 21287176 Review.
References
-
- Bent AF, Innes RW, Ecker JR, Staskawicz BJ (1992) Disease development in ethylene-insensitive Arabidopsis thaliana infected with virulent and avirulent Pseudomonas and Xanthomonas pathogens. Mol Plant Microbe Interact 5: 372–378 - PubMed
-
- Bonas U, Conradsstrauch J, Balbo I (1993) Resistance in tomato to Xanthomonas campestris pv. vesicatoria is determined by alleles of the pepper-specific avirulence gene avrbs3. Mol Gen Genet 238: 261–269 - PubMed
-
- Ciardi JA, Tieman DM, Jones JB, Klee HJ (2001) Reduced expression of the tomato ethylene receptor gene LeETR4 enhances the hypersensitive response to Xanthomonas campestris pv. vesicatoria. Mol Plant Microbe Interact 14: 487–495 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
