

CD70+ antigen-presenting cells control the proliferation and differentiation of T cells in the intestinal mucosa
- PMID: 15937486
- PMCID: PMC1444945
- DOI: 10.1038/ni1212
CD70+ antigen-presenting cells control the proliferation and differentiation of T cells in the intestinal mucosa
Abstract
One unresolved issue in gut immunity is how mucosal T lymphocytes are activated and which antigen-presenting cell (APC) is critical for the regulation of this process. We have identified a unique population of APCs that is exclusively localized in the lamina propria. These APCs constitutively expressed the costimulatory molecule CD70 and had antigen-presenting functions. After oral infection of mice with Listeria monocytogenes, proliferation and differentiation of antigen-specific T cells occurred in the gut mucosa in situ and blockade of CD70 costimulation abrogated the mucosal T cell proliferation and effector functions. Thus, a potent CD70-dependent stimulation via specialized tissue-specific APCs is required for the proliferation and differentiation of gut mucosal T cells after oral infection.
Figures
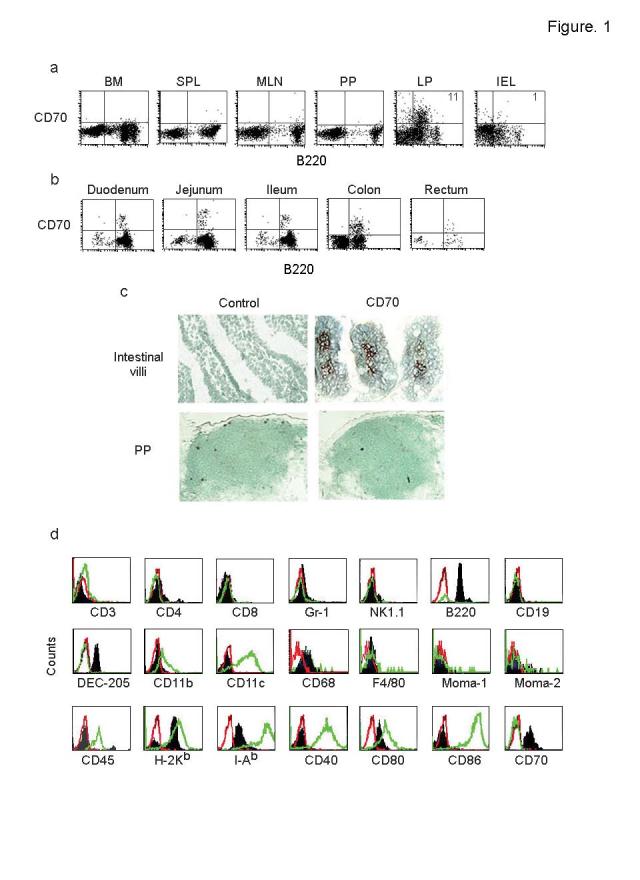
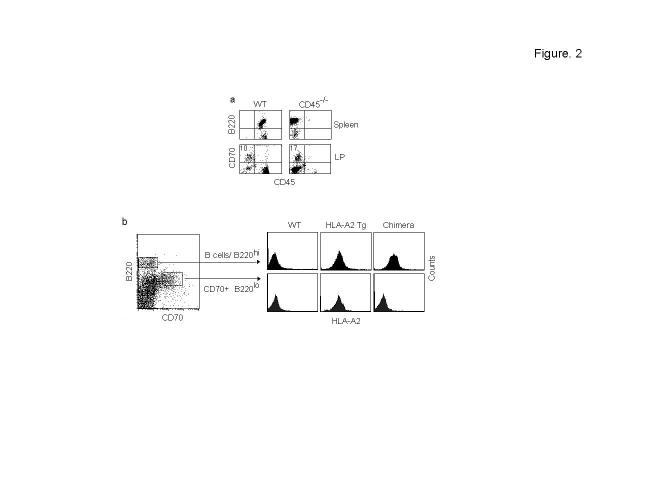
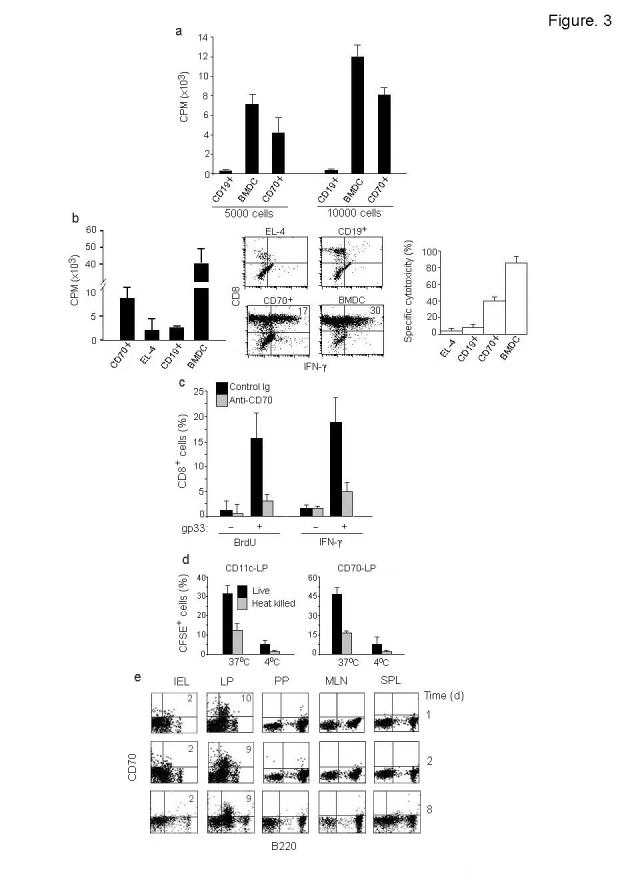
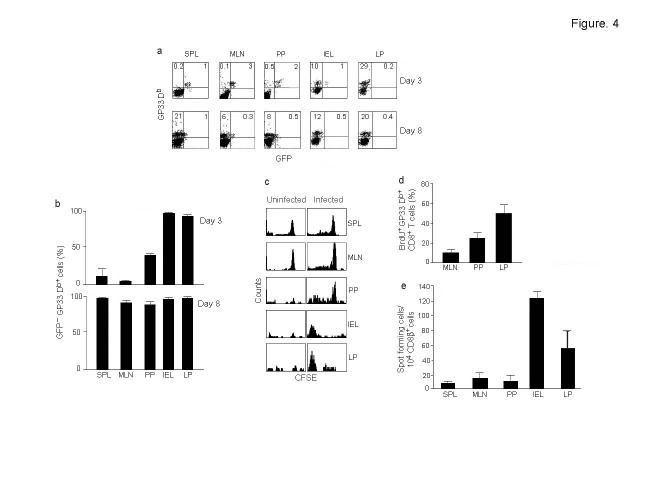
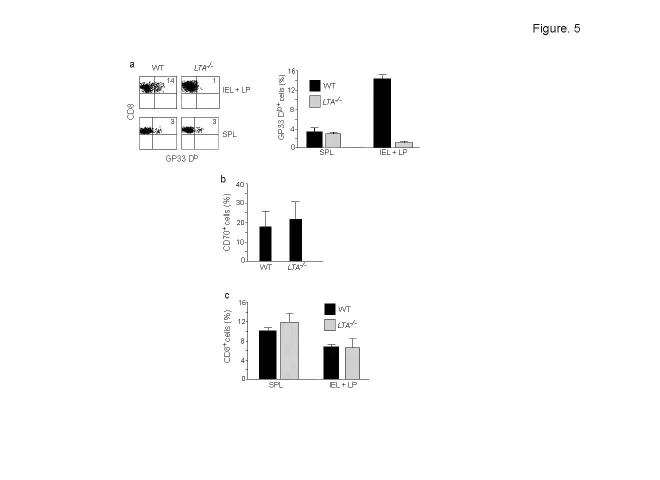
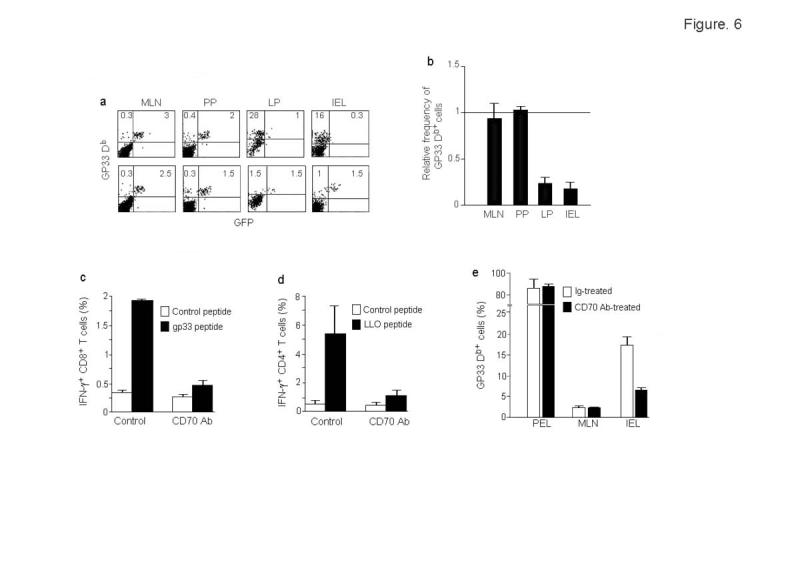
Comment in
-
Affecting the effectors: a kick in the gut?Nat Immunol. 2005 Jul;6(7):644-6. doi: 10.1038/ni0705-644. Nat Immunol. 2005. PMID: 15970934 No abstract available.
References
-
- Huleatt JW, Lefrancois L. Antigen-driven induction of CD11c on intestinal intraepithelial lymphocytes and CD8+ T cells in vivo. J Immunol. 1995;154:5684–93. - PubMed
-
- Beagley KW, Husband AJ. Intraepithelial lymphocytes: origins, distribution, and function. Crit Rev Immunol. 1998;18:237–54. - PubMed
-
- Pirzer UC, Schurmann G, Post S, Betzler M, Meuer SC. Differential responsiveness to CD3-Ti vs. CD2-dependent activation of human intestinal T lymphocytes. Eur J Immunol. 1990;20:2339–42. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
