

Methyl CpG-binding proteins induce large-scale chromatin reorganization during terminal differentiation
- PMID: 15939760
- PMCID: PMC2171616
- DOI: 10.1083/jcb.200502062
Methyl CpG-binding proteins induce large-scale chromatin reorganization during terminal differentiation
Abstract
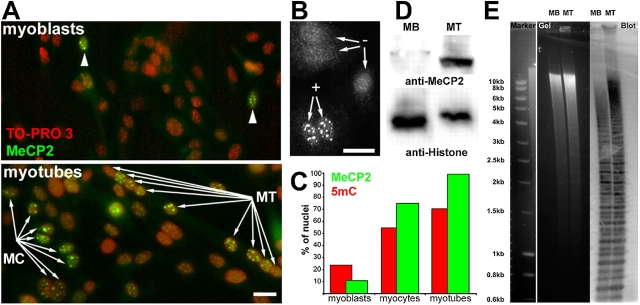
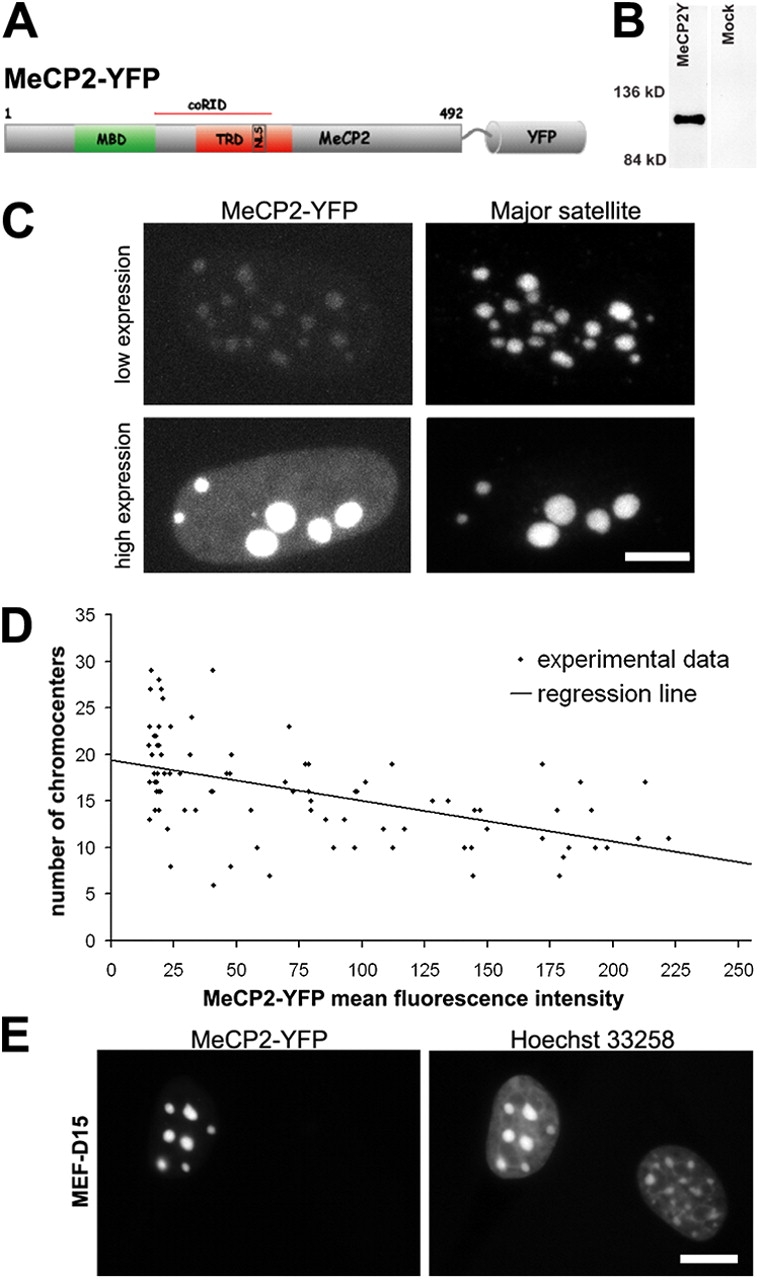
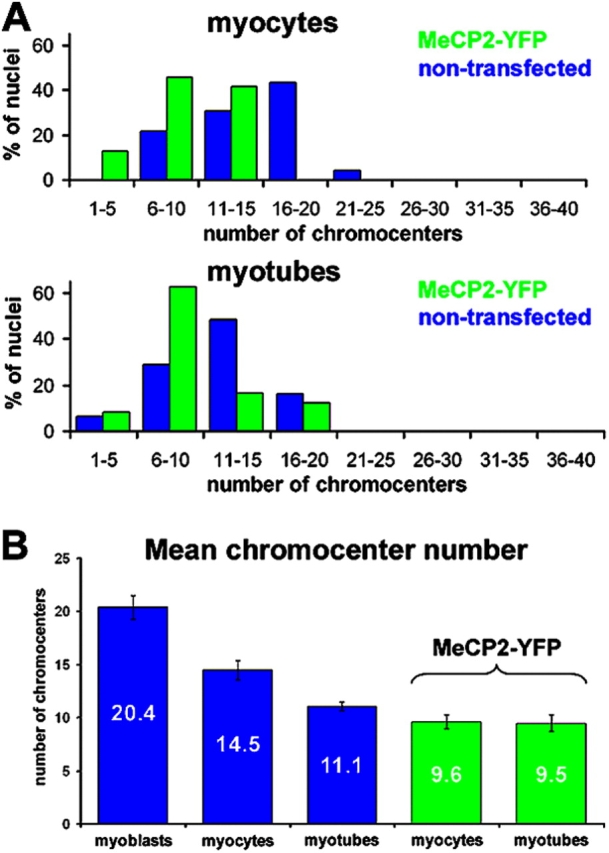
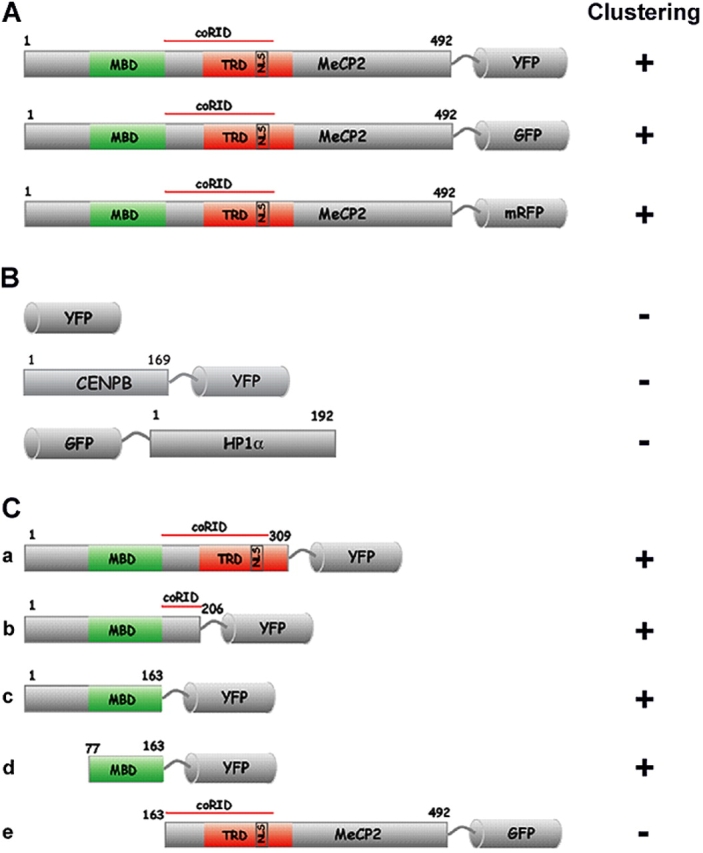
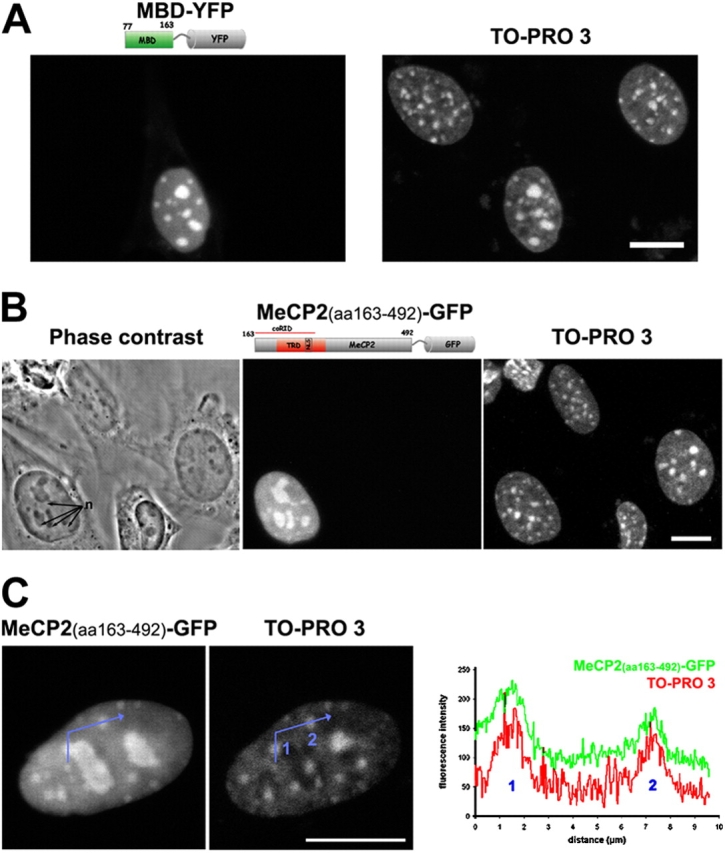
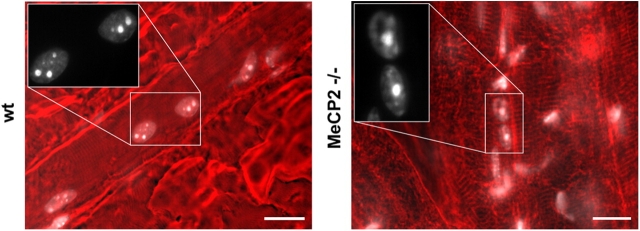
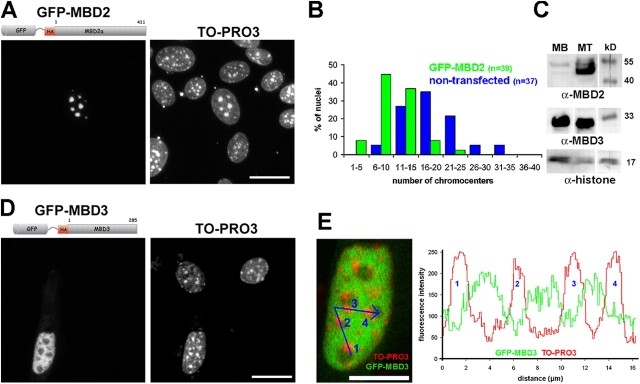
Pericentric heterochromatin plays an important role in epigenetic gene regulation. We show that pericentric heterochromatin aggregates during myogenic differentiation. This clustering leads to the formation of large chromocenters and correlates with increased levels of the methyl CpG-binding protein MeCP2 and pericentric DNA methylation. Ectopic expression of fluorescently tagged MeCP2 mimicked this effect, causing a dose-dependent clustering of chromocenters in the absence of differentiation. MeCP2-induced rearrangement of heterochromatin occurred throughout interphase, did not depend on the H3K9 histone methylation pathway, and required the methyl CpG-binding domain (MBD) only. Similar to MeCP2, another methyl CpG-binding protein, MBD2, also increased during myogenic differentiation and could induce clustering of pericentric regions, arguing for functional redundancy. This MeCP2- and MBD2-mediated chromatin reorganization may thus represent a molecular link between nuclear genome topology and the epigenetic maintenance of cellular differentiation.
Figures


References
-
- Amir, R.E., I.B. Van den Veyver, M. Wan, C.Q. Tran, U. Francke, and H.Y. Zoghbi. 1999. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nat. Genet. 23:185–188. - PubMed
-
- Amor, D.J., P. Kalitsis, H. Sumer, and K.H. Choo. 2004. Building the centromere: from foundation proteins to 3D organization. Trends Cell Biol. 14:359–368. - PubMed
-
- Beil, M., D. Durschmied, S. Paschke, B. Schreiner, U. Nolte, A. Bruel, and T. Irinopoulou. 2002. Spatial distribution patterns of interphase centromeres during retinoic acid-induced differentiation of promyelocytic leukemia cells. Cytometry. 47:217–225. - PubMed
-
- Bird, A.P., and A.P. Wolffe. 1999. Methylation-induced repression—belts, braces, and chromatin. Cell. 99:451–454. - PubMed
-
- Brown, K.E., S.S. Guest, S.T. Smale, K. Hahm, M. Merkenschlager, and A.G. Fisher. 1997. Association of transcriptionally silent genes with Ikaros complexes at centromeric heterochromatin. Cell. 91:845–854. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
