

Immune evasion versus recovery after acute hepatitis C virus infection from a shared source
- PMID: 15939788
- PMCID: PMC2213272
- DOI: 10.1084/jem.20042284
Immune evasion versus recovery after acute hepatitis C virus infection from a shared source
Abstract
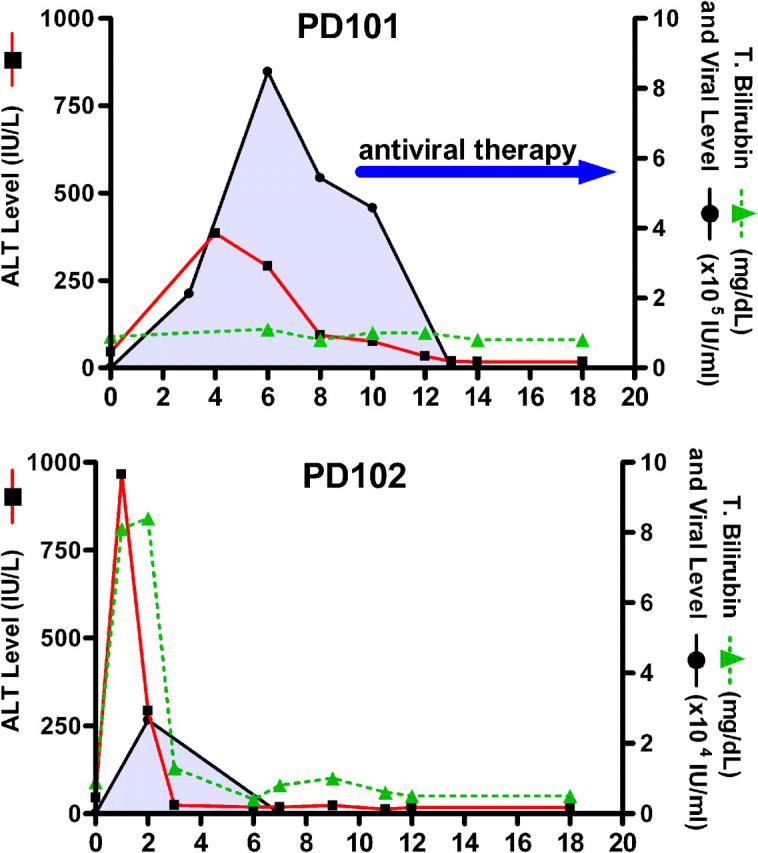
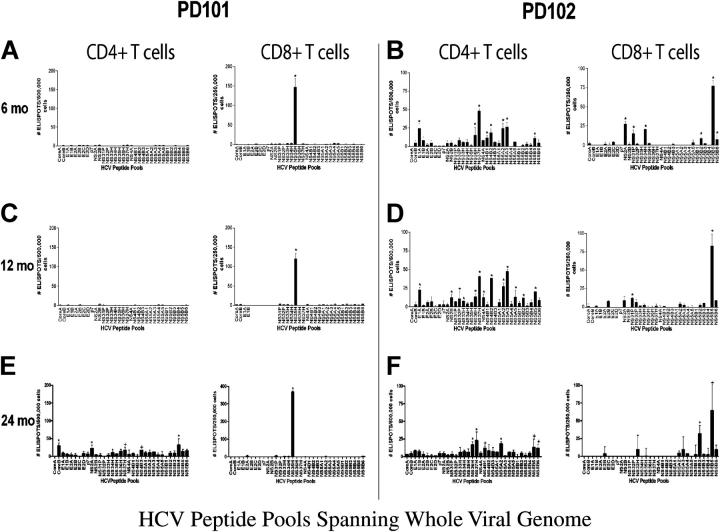
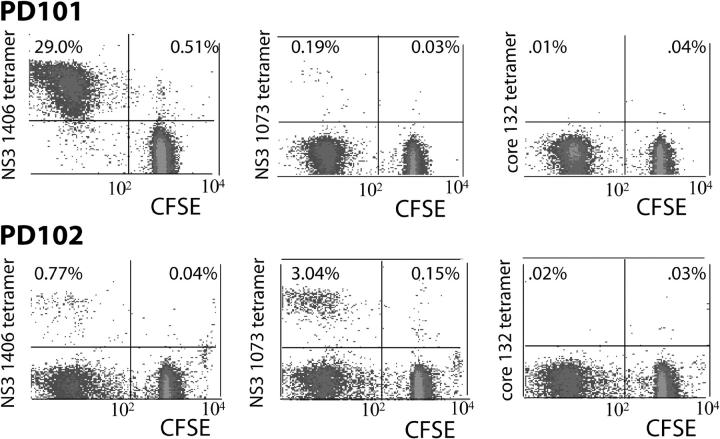
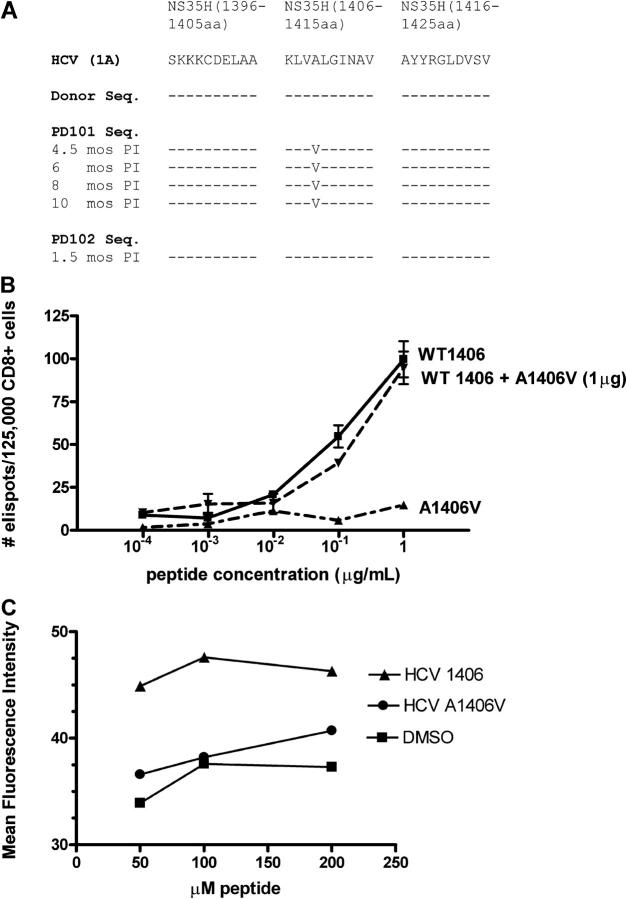
Acute infection with hepatitis C virus (HCV) rarely is identified, and hence, the determinants of spontaneous resolution versus chronicity remain incompletely understood. In particular, because of the retrospective nature and unknown source of infection in most human studies, direct evidence for emergence of escape mutations in immunodominant major histocompatibility complex class I-restricted epitopes leading to immune evasion is extremely limited. In two patients infected accidentally with an identical HCV strain but who developed divergent outcomes, the total lack of HCV-specific CD4+ T cells in conjunction with vigorous CD8+ T cells that targeted a single epitope in one patient was associated with mutational escape and viral persistence. Statistical evidence for positive Darwinian selective pressure against an immunodominant epitope is presented. Wild-type cytotoxic T lymphocytes persisted even after the cognate antigen was no longer present.
Figures

References
-
- Alter, M.J., 2002. Prevention of spread of hepatitis C. Hepatology. 36:S93–98. - PubMed
-
- Gordon, S.C. 2003. New insights into acute hepatitis C. Gastroenterology. 125:253–256. - PubMed
-
- Farci, P., A. Shimoda, A. Coiana, G. Diaz, G. Peddis, J.C. Melpolder, A. Strazzera, D.Y. Chien, S.J. Munoz, A. Balestrieri, et al. 2000. The outcome of acute hepatitis C predicted by the evolution of the viral quasispecies. Science. 288:339–344. - PubMed
-
- Cooper, S., A.L. Erickson, E.J. Adams, J. Kansopon, A.J. Weiner, D.Y. Chien, M. Houghton, P. Parham, and C.M. Walker. 1999. Analysis of a successful immune response against hepatitis C virus. Immunity. 10:439–449. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
