

Review
doi: 10.1038/sj.embor.7400441.
The effects of histone deacetylase inhibitors on heterochromatin: implications for anticancer therapy?
Affiliations
- PMID: 15940285
- PMCID: PMC1369099
- DOI: 10.1038/sj.embor.7400441
Item in Clipboard
Review
The effects of histone deacetylase inhibitors on heterochromatin: implications for anticancer therapy?
EMBO Rep.
2005 Jun.
Abstract
Histone acetylation regulates many chromosome functions, such as gene expression and chromosome segregation. Histone deacetylase inhibitors (HDACIs) induce growth arrest, differentiation and apoptosis of cancer cells ex vivo, as well as in vivo in tumour-bearing animal models, and are now undergoing clinical trials as anti-tumour agents. However, little attention has been paid to how HDACIs function in these biological settings and why different cells respond in different ways. Here, we discuss the consequences of inhibiting histone deacetylases in cycling versus non-cycling cells, in light of the dynamics of histone acetylation patterns with a specific emphasis on heterochromatic regions of the genome.
Figures
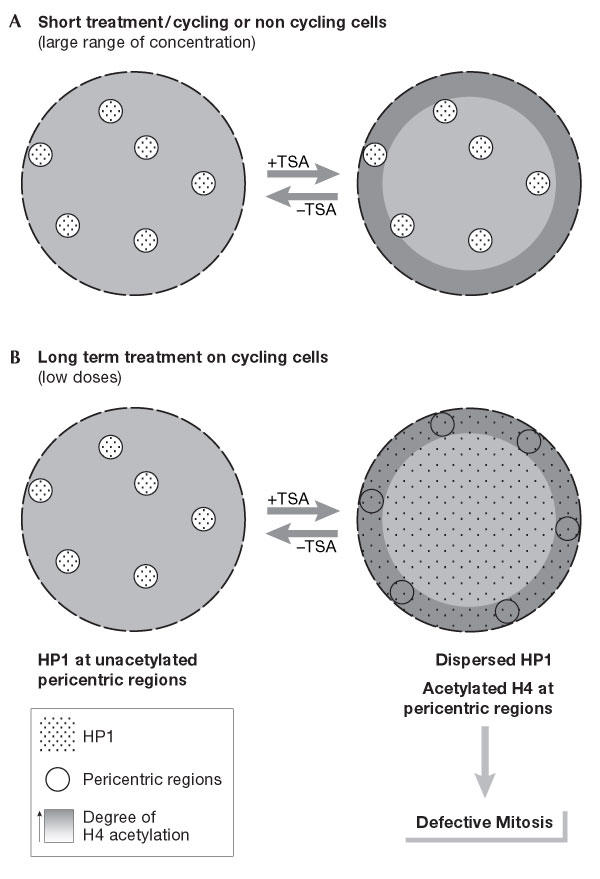
Reversible disruption of pericentric heterochromatin and centromere function by inhibiting deacetylases. (A) A very strong and rapid (within 90 min) enhancement of acetylated forms of H4 at the nuclear periphery is observed with a wide range of doses of histone deacetylase inhibitors (HDACIs) applied on a variety of cell types (Taddei et al, 1999). (B) Pericentric heterochromatin in cycling cells (immortalized human HeLa and U2OS, as well as mouse L929 cells) is specifically responsive to prolonged treatment with deacetylase inhibitors. At low doses, these defined regions relocate at the nuclear periphery and lose their properties of retaining heterochromatin protein 1 (HP1) proteins, which are spread throughout the nucleoplasm. Subsequent defects in chromosome segregation arise in mitosis. All these changes can reverse rapidly after drug removal (Taddei et al, 2001).
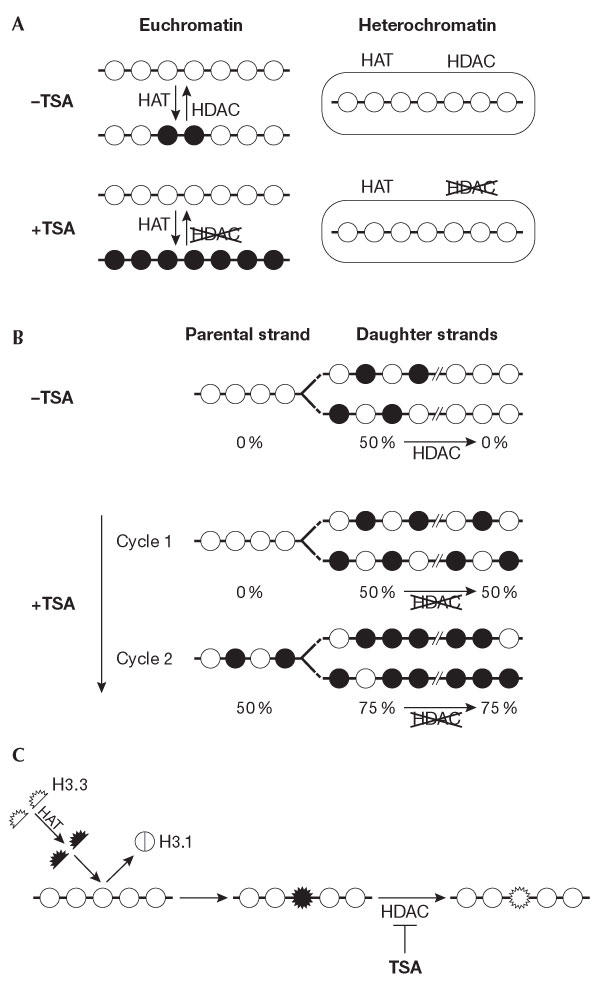
Different dynamics of histone acetylation. (A) Replication-independent dynamics. The acetylation state of a chromatin locus results from the antagonist activities of histone acetyltransferases (HATs) and histone deacetylases (HDACs) on pre-existing nucleosomes. These two activities are locally regulated. Indeed, inhibiting deacetylases by trichostatin A (TSA) stabilizes the acetylation state of histone H4 in euchromatin regions with a very strong efficiency at the nuclear periphery, whereas it has no effect on heterochromatin regions (Taddei et al, 2001). (B) Replication-dependent dynamics of histone acetylation. After DNA replication, DNA daughter strands are rapidly wrapped into nucleosomes that contain a mixture of parental and newly synthesized histones. The bulk of newly synthesized histone H4 is acetylated at lysines 5 and 12 by a cytoplasmic HAT and is incorporated into new chromatin in this form. During late S phase, the acetates at H4 lysines 5 and 12 are rapidly lost in heterochromatin regions, probably owing to the presence of an HDAC at the site of chromatin assembly that can be inhibited by TSA treatment. This inhibition will lead to the maintenance of the acetylated state of newly incorporated histones, which dilutes by twofold the non-acetylated parental histones at each round of replication in the presence of TSA (Taddei et al, 1999). (C) Dynamics of histone acetylation by histone exchange. The variant H3.3 (expressed throughout the cell cycle) can be deposited into chromatin at any time. It has been shown that H3.3 is enriched in modifications that are associated with transcriptional activity. We propose that it can be acetylated before incorporation, thus providing another means to change chromatin patterns. The action of HDACIs that is envisaged in this context is depicted.
References
-
- Almouzni G, Khochbin S, Dimitrov S, Wolffe AP (1994) Histone acetylation influences both gene expression and development of Xenopus laevis. Dev Biol 165: 654–669 - PubMed
-
- Craig JM, Earle E, Canham P, Wong LH, Anderson M, Choo KH (2003) Analysis of mammalian proteins involved in chromatin modification reveals new metaphase centromeric proteins and distinct chromosomal distribution patterns. Hum Mol Genet 12: 3109–3121 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
