

The exchangeability of amino acids in proteins
- PMID: 15944362
- PMCID: PMC1449787
- DOI: 10.1534/genetics.104.039107
The exchangeability of amino acids in proteins
Abstract
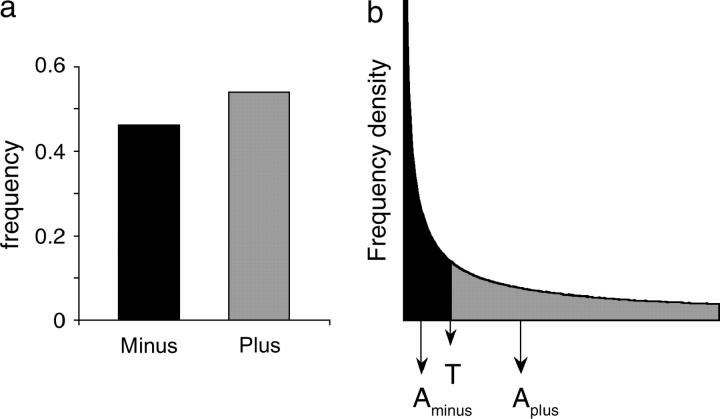
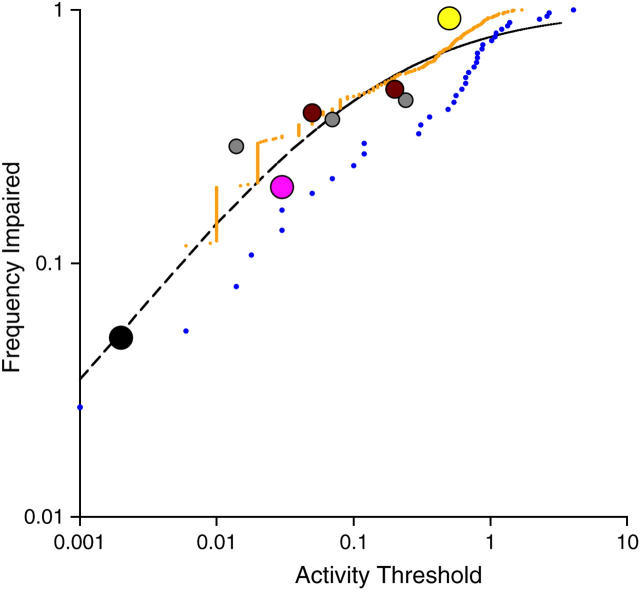
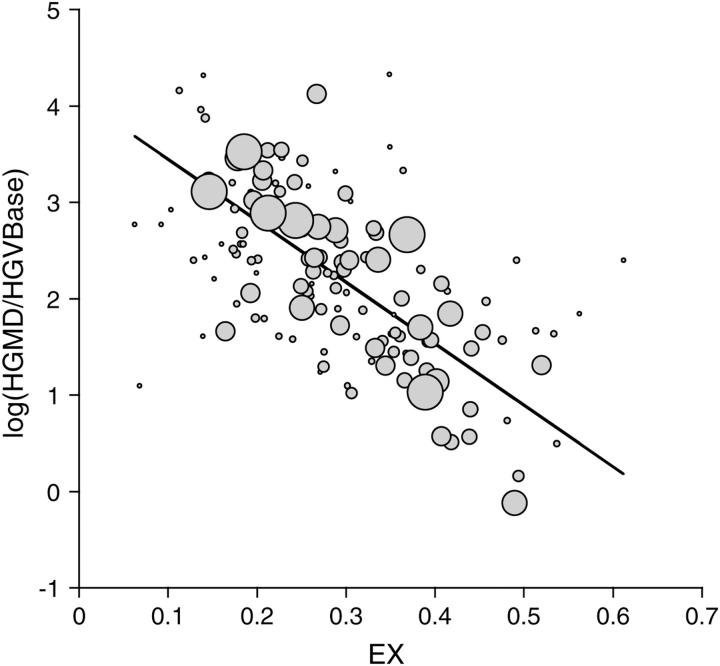
The comparative analysis of protein sequences depends crucially on measures of amino acid similarity or distance. Many such measures exist, yet it is not known how well these measures reflect the operational exchangeability of amino acids in proteins, since most are derived by methods that confound a variety of effects, including effects of mutation. In pursuit of a pure measure of exchangeability, we present (1) a compilation of data on the effects of 9671 amino acid exchanges engineered and assayed in a set of 12 proteins; (2) a statistical procedure to combine results from diverse assays of exchange effects; (3) a matrix of "experimental exchangeability" values EX(ij) derived from applying this procedure to the compiled data; and (4) a set of three tests designed to evaluate the power of an exchangeability measure to (i) predict the effects of amino acid exchanges in the laboratory, (ii) account for the disease-causing potential of missense mutations in the human population, and (iii) model the probability of fixation of missense mutations in evolution. EX not only captures useful information on exchangeability while remaining free of other effects, but also outperforms all measures tested except for the best-performing alignment scoring matrix, which is comparable in performance.
Figures
References
-
- Atchley, W. R., T. Lokot, K. Wollenberg, A. Dress and H. Ragg, 2001. Phylogenetic analyses of amino acid variation in the serpin proteins. Mol. Biol. Evol. 18: 1502–1511. - PubMed
-
- Axe, D. D., N. W. Foster and A. R. Fersht, 1998. A search for single substitutions that eliminate enzymatic function in a bacterial ribonuclease. Biochemistry 37: 7157–7166. - PubMed
-
- Benner, S. A., M. A. Cohen and G. H. Gonnet, 1994. Amino acid substitution during functionally constrained divergent evolution of protein sequences. Protein Eng. 7: 1323–1332. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
